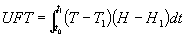Octubre 2013
![]()
![]()
Octubre 2013
ADAPTACIONES Y POSIBLES MANIPULACIONES MICROCLIMÁTICAS
A FAVOR DE UNA MAYOR PRODUCCIÓN AGRÍCOLA
Walter Ritter Ortiz
Centro de Ciencias de la Atmósfera, UNAM. Circuito exterior s/n, Ciudad Universitaria, Deleg. Coyoacan, México, D. F.
email:
walter@atmosfera.unam.mx
INTRODUCCIÓN
Frecuentemente las interacciones de un organismo vivo con su ambiente son tan complejas que solamente una relación estadística puede esperarse a ser utilizada.
Es conveniente considerar el "stress" ambiental de corta duración (minutos a días) separadamente de las influencias a largo plazo (meses a años).
Los organismos no siempre son capaces de aclimatarse al "stress" de largo plazo, sin embargo puede tomar muchos años antes de que un efecto de daño pueda ser detectado.
El microclima es cambiado en forma considerable por el hombre, por la vegetación y en ocasiones por los animales e insectos.
Menos dramático que el hombre y los animales, pero más significativo, es el efecto de la vegetación sobre la temperatura, humedad y viento cerca del suelo.
La vegetación a menudo causa que el subsuelo permanezca frío y anegado, sin embargo la temperatura del aire dentro de la cubierta vegetativa es mayor que sobre la tierra.
El sombreado artificial de una plantación cambia no solamente el balance de radiación, sino también la temperatura de la hoja, la razón de transpiración y la humedad del suelo.
Así también una población animal puede deliberadamente introducirse en un área cerrada en estudios de productividad, generando una larga cadena de reacciones ambientales:
El sobre pastoreo modifica el microclima, el cual a su vez disminuye el abastecimiento de alimento.
En algunos casos la respuesta secundaria del ecosistema puede ser de interés primordial.
Las plantas a menudo tienen amplias zonas de confort y muchas de sus necesidades ambientales (agua y luz) están en gran abundancia.
La sobrevivencia y crecimiento dependen entonces de unos pocos factores limitantes.
Estos factores deben ser identificados y las fronteras de su "stress" determinarse, llamando "climatología ecológica" a la encargada del estudio de las relaciones entre el clima y la vegetación natural, así como con los insectos y distribución de otros animales (Munn, 1970).
Las oscilaciones de fronteras ecológicas señalan una complejidad y laboriosidad de determinación, requiriendo un monitoreo de muchas estaciones; además en el caso de vegetación, el movimiento de fronteras puede depender de eventos raros como heladas en primavera, sequías, etc.
En la parte central del bosque, hay poca variación en el crecimiento vegetativo de año a año, existiendo un equilibrio ambiental creando el bosque su propio microclima favorable.
Los árboles de la orilla son más sensitivos y pueden ser usados como índices de variación climática.
El determinismo climático considera que las formas de vida conocida, son el producto o resultante del ambiente.
El "stress" ambiental actuando durante muchos años o centurias, conduce a una aclimatización con cambios permanentes en la fisiología o el comportamiento.
En principio: suelo y vegetación deben ser un espejo de las condiciones climáticas normales.
La búsqueda de relaciones precisas entre ellos es uno de los más importantes desafíos de la climatología moderna.
Los ecólogos están interesados en determinar si la vegetación está en equilibrio con su ambiente y si las comunidades existentes son más eficientes, en términos de productividad y uso del agua.
Un climatólogo experimentado puede estimar las características climáticas regionales, desde un conocimiento de la naturaleza del suelo, la orografía, las propiedades térmicas, disponibilidad de humedad y cubierta vegetal.
Las plantas al brotar del suelo, pueden experimentar en un sólo día rangos de temperatura, que para el hombre y otras formas de vida, corresponderían a cambios del Trópico al Ártico.
Los gradientes microclimáticos de temperatura, humedad y viento son mayores en la interfase del suelo.
Cuando el sol está brillando, el aire dentro de los primeros milímetros del suelo, puede ser hasta de 10°C más caliente que a una altura de 3 metros.
En cambio, en las noches claras y tranquilas, un gradiente inverso de igual magnitud puede darse.
En una escala menor, grandes gradientes de temperatura existen en el aire adyacente a las hojas iluminadas, así cuando el viento es ligero, dinámicas de circulación local pueden desarrollarse.
Las hojas también regulan su abastecimiento de calor por enfriamiento del agua que evaporan al transpirar.
Algunas plantas cambian su orientación para presentar un mínimo de superficie a la radiación.
Otro "stress" en las plantas es la disminución de la humedad del suelo; la planta se ajusta a ésta cerrando sus estomas (reduciendo su pérdida de humedad por transpiración) y extendiendo sus raíces a mayores profundidades.
En días soleados, en zonas de climas secos, algunas especies disminuyen su evapotranspiración después del medio día debido a un "stress" temporal de humedad tendiendo a la recuperación por la tarde.
Los bosques tienen un efecto significativo en la humedad del suelo, el nivel freático y el tiempo e intensidad de los escurrimientos del agua.
Excepto cerca de la superficie, el suelo de un bosque es menos húmedo que el de un pastizal, debido a la profunda penetración de las raíces y la gran área de hojas transpirando (un árbol de maple tiene un área de hojas transpirando de aproximadamente 4000 mt2.).
Para una superficie uniforme, si las condiciones meteorológicas no cambian, la razón de evaporación varía conforme al producto de la velocidad del viento y el gradiente vertical de presión de vapor.
Los vientos y el gradiente de vapor de presión de humedad tienen un ciclo diurno y por lo tanto están correlacionados.
La caña de azúcar en zonas irrigadas del desierto pierde cinco veces más humedad por transpiración que un lago sin vegetación.
Las condiciones micrometeorológicas son relativamente constantes al medio día y media noche, pero cambian rápidamente al amanecer y oscurecer.
Oscilaciones de las fronteras ecológicas señalan la necesidad de monitoreo sobre muchas estaciones, donde en algunos experimentos micrometeorológicos, hasta 100 medidas por segundo son requeridas en un sensor.
Muchos tipos de respuestas ecológicas parecen ser del tipo log-normal, parametrizada por una sola variable en muchas ocasiones, conocida como TL50 (nivel de tolerancia del 50%), Munn (1970) ajustándose a una hipérbola de la forma
En lo que
a y b son constantes empíricas y t es el tiempo de exposición.El crecimiento de las plantas, deben ser relacionados no a fechas del calendario, sino a eventos fenológicos, así el efecto de anomalías de la humedad y temperatura, pueden ser relacionadas específicamente a cada estado de desarrollo del cultivo.
Una cubierta vegetal es una fuente de CO2 en la noche, pero un sumidero en el día.
El flujo de CO2 requerido en la fotosíntesis, es mucho mayor que la liberada por respiración aunque a temperaturas mayores a 35°C es válido lo contrario donde por cada gramo de CO2 asimilado, se necesitan aproximadamente 90 gramos de vapor de agua transpirada, Lowry (1970)
En la fotosíntesis, las hojas de varias especies requieren hasta 4 minutos para responder a un incremento en la intensidad de la luz, pero en menos de 30 segundos reaccionan a una disminución.
Monteith (1965) ha establecido la relación:
entre fotosíntesis e iluminación.
Donde "a" es una resistencia multiplicada por un índice volumétrico de densidad de cosecha; y "b" es inversamente proporcional a la eficiencia cuántica de la fotosíntesis en luz débil.
Las razones de respiración y fotosíntesis dependen de la temperatura.
En este último caso la adaptación climática ha demostrado ser importante; La temperatura óptima es de aproximadamente 12°C en regiones alpinas, 20°C en zonas templadas y en 30°C en las tropicales.
La respuesta fisiológica de las plantas además de la temperatura depende de las horas luz.
La floración y producción del fruto es a menudo críticamente dependiente de la longitud del día o fotoperíodo y no de la intensidad de la luz por lo que las plantas de fotoperíodo largo no florean o no se pueden reproducir en los trópicos.
En comparaciones de la razón de crecimiento de cultivos agrícolas, el número de horas de insolación es muy importante y se ha sugerido que la unidad fototérmica (UFT) (°C/hora) sea normalizada a una longitud estandard del día, teniéndose que:
Con
T1 y H1 como la temperatura y horas de "insolación de referencia".A menudo las investigaciones agrícolas describen en detalle los tipos de fertilizantes y frecuencias de irrigación que deben usarse, pero fallan en mencionar si existe uniformidad microclimática.
Así, dos campos de cultivo que pueden parecer iguales, pueden de hecho ser muy diferentes.
Muchos métodos estadísticos contienen consideraciones que raramente son satisfechas en biometeorología (aleatoriedad y linealidad).
Los métodos estadísticos son útiles en determinar si existe una asociación entre dos o más variables, pero sin que se puedan hacer inferencias acerca de causa y efecto; así por ejemplo las razones de fotosíntesis y transpiración están correlacionadas porque cada una está correlacionada con otros factores ambientales tales como el flujo de radiación solar, temperatura y deficiencia de nitrógeno en el suelo.
El análisis de varianza, cuadrados latinos, Ji-cuadrada y las -pruebas t y F ayudan a decidir si se acepta en la presencia de ruido experimental, la realidad de situaciones de similaridad, diferencias o de asociaciones.
CLIMAS Y COMPORTAMIENTO
La ciencia ha estudiado la relación entre el estado del tiempo y una diversidad de actividades humanas, como podría ser la frecuencia de casos de asalto y agresión, a la conducta de los niños en clase y casos de suicidio.
El efecto más pronunciado del clima sobre el hombre según Munn (1970), es el que ejerce en su vida personal, a través de su influencia sobre el cuerpo, mente y emociones.
En muchos casos la relación es obvia y claramente biológica.
En otros, el tiempo puede influir en el hombre de un modo mucho más complejo, oscuro e indirecto.
Los estados de mal humor y la presión atmosférica baja, están asociados.
En los días en que la humedad es baja y el barómetro sube, las personas tienden a realizar su trabajo alegremente, funcionan con más eficiencia y contemplan la vida con optimismo.
Pero las mismas personas en un día bochornoso, regañan más a sus hijos, están violentos con sus compañeros y se sienten taciturnos y sombríos además de pesimistas.
En circunstancias normales la mayor parte de las personas soportan estas dificultades inducidas por el tiempo, pues la perspectiva y el sentido del humor les ayudan; pero para las personas cuya condición física está por debajo de lo normal, los efectos opresivos del mal tiempo pueden hacer que una salud mediana se convierta en seria enfermedad.
Y en casos de personas que están sujetas a tensiones emocionales, puede ser determinante de repentinos actos de agresión.
La temperatura, más que cualquier otra circunstancia afecta los estados emotivos que conducen a la pelea.
Desde hace tiempo se atribuye también una profunda influencia sobre las personas a los vientos que soplan con regularidad y monótonamente durante semanas seguidas.
El tiempo más sano para la mayoría de las personas no es necesariamente el buen tiempo.
Algunos médicos creen que cualquier estado del tiempo, bueno o malo, puede tener efectos perjudiciales si persiste demasiado.
En primer lugar, es deprimente por ser tan monótono, pero es también perjudicial físicamente; el cuerpo se hace menos adaptable a los cambios y es por lo tanto, más vulnerable a ellos cuando se presentan.
Pero también el tiempo muy variable puede ser perturbador, así el más importante agente de repentinos cambios de tiempo, la tormenta ciclónica, parece desencadenar profundas perturbaciones en cuerpos y mentes.
A medida que se acerca el frente tormentoso y el barómetro desciende y la humedad aumenta, la gente se siente incomprensiblemente perturbada por una sensación de incapacidad para alcanzar su eficiencia mental normal, o para realizar "tareas difíciles".
Los niños se vuelven irritables y los adultos pendencieros.
Al pasar el frente tormentoso, el tiempo y las emociones varían radicalmente, la humedad disminuye, la presión barométrica aumenta, el aire se vuelve fresco, transparente y vigorizador, la gente está de buen humor y se realizan difíciles tareas con facilidad.
Si es cierto que las variaciones de presión atmosférica afectan nuestros pensamientos y sentimientos, ¿cómo ocurre? La ciencia ignora la respuesta, pero algunos opinan que la clave puede estar en los iones, pequeñas partículas de materia con carga eléctrica que existe en la atmósfera.
Generalmente se encuentran en el aire a razón de cinco positivos por cuatro negativos.
Los iones negativos se componen parcialmente de oxígeno, que puede ser beneficioso para el cuerpo humano; los iones positivos están en parte compuestos por CO2 que puede ser perjudicial.
Es significativo que entre los factores que se sabe que producen un desequilibrio de iones positivos se encuentra el "smog" y ciertas clases de calefacción y de equipos de acondicionamiento del aire.
En experimentos sobre voluntarios que fueron expuestos a corrientes de iones negativos, demostraron que sus reacciones visuales habían mejorado y que podrían trabajar mucho más sin mostrar fatiga, haciendo que el sistema respiratorio del cuerpo funcionase mejor.
Se ha sugerido que diversas circunstancias de la atmósfera misma pueden producir un desequilibrio de iones; quizá inmediatamente antes de una tempestad el aire está sobrecargado de iones positivos y por lo tanto, de dióxido de carbono; en tanto el equilibrio se desplaza en el sentido opuesto cuando estalla la tormenta, descargando un exceso de iones negativos y de oxígeno al aire.
El dióxido de carbono podría explicar los efectos biológicos deprimentes y el oxígeno la euforia.
MICROCLIMAS EN ANIMALES
La interacción entre la atmósfera y los animales representan uno de los niveles de mayor complejidad en la capa frontera.
El balance energético de un animal, puede escribirse según Oke (1970) como:
Donde Qm es la razón de producción de calor por procesos metabólicos y QS es el cambio neto de almacenamiento de calor en el cuerpo.
En este balance: Qm es siempre una fuente de calor. Q* es también una fuente de calor, especialmente durante períodos de fuerte radiación; y QH y QG pueden ser fuentes de calor si el aire que rodea al animal o el suelo con el que está en contacto, es más caliente que la temperatura del cuerpo del animal.
Tanto Q*, QH, QE, como QG, todos representan canales de pérdida de calor para disipar el calor metabólico producido por el animal.
El almacenamiento neto de calor, puede ser una ganancia o pérdida de energía, pero en muchos animales debe permanecer próximo a cero, porque el rango tolerable de temperaturas del cuerpo es pequeño a menos que el animal esté en una madriguera o inmerso en agua la ganancia o pérdida de calor por conducción es usualmente despreciable comparado con otros medios de intercambio.
Si consideramos el intercambio de radiación en onda corta durante el día, algunas áreas del cuerpo estarán iluminadas recibiendo radiación directa y difusa, mientras otras estarán sombreadas, recibiendo sólo radiación difusa.
Las áreas iluminadas toman diferentes formas, dependiendo de la geometría del animal y de la luz incidente.
Además la parte inferior del animal recibirá más de la energía reflejada del suelo.
El albedo del animal no es espacialmente uniforme. Si agregamos el hecho de que el animal es capaz de moverse a diferentes ambientes, la situación se vuelve excesivamente complicada y por lo tanto difícil de monitorear o modelar.
Las pérdidas por radiación en onda larga y calor sensible, dependen de la temperatura superficial y ésta, puede ser mayor donde el pelaje es delgado o ausente (orejas, abdomen, cola, etc.).
Así también en animales que sudan el enfriamiento por evaporación puede estar enteramente localizado en la tráquea respiratoria.
El correspondiente balance volumétrico de agua en un animal está dado por:
Donde: BV es la entrada neta de agua (usualmente con la comida y bebida, aunque algunos animales son capaces de absorber agua a través de su cubierta exterior); U es el agua perdida por orina y heces y Ah es el cambio de almacenamiento de agua en el cuerpo (para animales pequeños debe igualarse directamente al cambio de peso).
Las pérdidas por evaporación (Ea) pueden realizarse por tres vías:
- El agua puede evaporarse y difundirse directamente a través de la piel.
- Puede liberarse a través de los poros en la piel por las glándulas sudoríperas y ser evaporada de la superficie de la piel.
- La evaporación puede ocurrir de las superficies húmedas de la tráquea respiratoria.
El ser vivo depende de diferentes reacciones químicas en su actividad diaria y estas reacciones químicas son cuidadosamente coordinadas, donde cada proceso se da en su razón justa.
Todas las reacciones químicas se aceleran conforme la temperatura aumenta, pero a diferentes velocidades; así que un grupo de reacciones que fueran propiamente coordinadas a una temperatura, no lo serán con otra.
A más simplicidad del organismo más simple su química; esperaremos que los organismos más simples sean menos afectados por cambios de temperatura que los organismos más complejos.
Esto es verdad para algunas bacterias que pueden vivir sobre un rango de temperaturas que excedan los 100°F, mientras que para el hombre un cambio de sólo unos pocos grados puede ser de consideraciones graves.
El calor generado por los organismos tiende a calentarlos y a cambiar su balance químico.
Varias formas han desarrollado los organismos para liberarse de su calor: pueden hacerlo por conducción (muy importante en organismos marinos) o radiación en la forma de radiación en onda larga o infrarroja.
La razón a la que los organismos pierden calor debe de estar en balance con la razón en que lo generan; si la pérdida es muy pequeña el organismo se calienta y si la pérdida es muy grande, el organismo se enfría.
El buen funcionamiento de un organismo vivo depende de la relación que tenga con su medio ambiente.
Existen dos tipos básicos de organismos que se distinguen por el grado de control interno de su balance térmico con el ambiente exterior.
Los organismos cuya temperatura está totalmente determinada por el ambiente que lo rodea, plantas y animales de sangre fría (insectos, reptiles, peces) pertenecen a este grupo.
Por otra parte, otros son capaces de mantener una temperatura del cuerpo relativamente constante por medio de mecanismos fisiológicos que varían la producción del calor metabólico o la pérdida de calor por radiación, conducción y convección. Los animales de "sangre caliente" pertenecen a este grupo.
Grandes variaciones de las condiciones normales (cuando QS se hace significativo y la temperatura sube o baja demasiado) es perjudicial para el animal.
Los animales de "sangre fría" no gastan energía en termoregulación, su razón metabólica está determinada solamente por su temperatura.
Los animales de "sangre caliente" para mantener su independencia de la temperatura ambiental, tienen que pagar un enorme costo energético.
Su razón metabólica debe incrementar linealmente con la disminución de la temperatura ambiental, porque a más baja la temperatura externa, más trabajo se debe de hacer para mantener su temperatura constante.
Sin embargo, arriba de un límite de temperatura, el animal puede permanecer en forma confortable con un mínimo relativamente constante de metabolismo, lo cual es llamada "la zona de mínimo metabolismo".
Para mayores temperaturas la razón metabólica aumenta en forma no-lineal.
Los animales de "sangre caliente" han desarrollado respuestas especiales al stress ambiental.
Estas incluyen cambios en el metabolismo, dilatación y contracción de los vasos sanguíneos, aumento o decremento del pulso, sudado y erección de pelo o plumas.
El calor metabólico producido depende del estado del ambiente físico circundante, así como de la dieta, tamaño del cuerpo, edad y nivel de actividad del animal, aumentando su actividad basal con el tamaño y disminuyendo con la edad.
Los requerimientos energéticos son proporcionales al área de la superficie del animal, pero solamente a un exponente de dos tercios de su masa.
Así, un animal grande, requiere relativamente poca energía para conservar la temperatura de su cuerpo estable; mientras que un animal pequeño requiere más energía por unidad de masa.
Un elefante necesita 33J Kg-1 y un ratón 837J Kg-1.
La razón de la masa del animal al área de su superficie es muy importante.
Aquéllos con pequeñas razones masa-superficie (Delgados, piernas largas) son buenos para intercambiar calor y están bien adaptados a la disipación de calor y ambientes cálidos.
Aquéllos con grandes razones de superficie-masa están mejor adaptados a la conservación del calor y para vivir en ambientes fríos.
Los apéndices de los animales, usualmente tienen pequeñas razones masa superficie en comparación con el resto del cuerpo y así, son buenos sitios de intercambio de calor.
-INSECTOS
Aunque existe un gran número de especies de insectos, todos tienen la característica general de ser pequeños, y que sus razones masa-superficie es también pequeña y al igual que las hojas, son buenos intercambiadores, pero malos almacenadores de energía.
Esto señala que los insectos dependen fuertemente de las condiciones térmicas que los rodean.
Sin embargo, ellos no tienen una fuente externa de continuo abastecimiento de agua para mantener sus razones de evaporación.
El agua de su cuerpo la obtienen a través de su alimento, bebida y en algunos casos absorción de vapor de agua.
La pierden por evaporación de su cubierta exterior, su sistema respiratorio y por excreción.
Los animales pequeños no pueden permitirse el lujo de una evaporación libre sin una rápida disecación, así que los insectos se caracterizan por un exterior relativamente impermeable, siendo QE despreciable en su balance energético, (Oke, 1978).
Durante el día cuando el insecto está expuesto al sol y está activo, la ganancia neta de calor vía radiación (Q*), producción de calor metabólico (Qm) y conducción (Qp) puede hacerse excesivo así que Q* aumenta y el insecto se aproxima a su límite superior de temperatura para su actividad o sobrevivencia.
Como no tiene pérdidas por evaporación, el único canal posible de liberación de calor es a través del calor sensible (QH).
El comportamiento para evitar el frío incluye un amplio rango de actitudes.
Por ejemplo, algunas mariposas orientan sus alas y cuerpos para: obtener el máximo de intensidad de radiación solar o se pegan junto a superficies calientes y restringen su vuelo a un metro de altura para minimizar las pérdidas convectivas.
Estas técnicas permiten a algunas especies elevar su temperatura hasta en 17°C arriba de la del aire y permanecer activas aún cuando la temperatura del aire sea baja.
Algunos insectos tienen un rango restringido de temperaturas del cuerpo, dentro del cual es posible volar.
Sin embargo, son capaces de elevar la temperatura de su cuerpo por generación muy rápida de su calor metabólico.
En esta actividad, los músculos para volar son operados en forma vigorosa, lo cual permite producir un metabolismo muchas veces superior que cuando está en reposo y este calor es conservado para su posterior uso en el vuelo.
Con respecto al balance de agua en animales, la cantidad de disponibilidad de agua depende de su masa, pero la razón de evaporación depende del área de su superficie.
Así para animales pequeños con pequeñas razones masa-superficie, el enfriamiento por evaporación es un lujo imposible de tener, ya que si éste se diera se daría rápidamente una deshidratación.
Los animales grandes pueden usar el agua para su termorregulación y pueden sobrevivir mayor tiempo sin recurrir a ella.
ÍNDICES DE CONFORT HUMANO
La zona de confort es bastante incierta y no es fácil de especificar, o de explicar, ya que depende de factores como:
a) La producción de calor metabólico,
b) El intercambio radiativo con el ambiente y el Sol,
c) La pérdida calorífica por procesos convectivos,
d) Conducción calorífica al suelo y otros objetos,
e) Pérdida de calor por los pulmones,
f) Efecto de la ropa para reducir la pérdida por conducción,
convectiva, radiativa y conductiva,
g) Variables fisiológicas, como el sexo, edad, salud, hora de
ingestión de alimentos y hora del día,
h) Efectos fisiológicos como longitud de la oscuridad, lluvia,
nevadas y estado general de la mente.
Además, en general, conforme el balance de calor se aleja de lo normal, el stress incrementa.
Se considera que una temperatura media anual de 14°C es un valor central razonable para el control humano y para la actividad biológica.
Con humedad relativa muy baja, los labios se secan y la piel empieza a escocer.
Conforme el aire se calienta, los cambios de humedad relativa se vuelven más importantes en la sensación de confort.
El efecto de un cambio en la humedad relativa puede ser modelada por la ecuación:
= PC CN (qs qa ) / (ts- ta)
Donde O < PC < 1 es el índice de permeabilidad de humedad (ropa usada) que es independiente de la humedad y temperatura y Ch
γ v/γh la resistencia al transporte de vapor de agua y la resistencia de transporte por calor convectivo ts y qs es la temperatura y humedad específica de la piel.Así también se ha usado la "entalpia" o contenido total de calor de la atmósfera excediendo los -18°C para aire seco y 0°C para el vapor de agua, (Munn, 1970) con la ventaja de que sus dimensiones (kcal/kg) son consistentes con las unidades metabólicas (KCal/kg-hr )
i = Cp (Tw+ 1.555 Pa/e*)
Si la temperatura del aire que nos rodea, así como paredes y suelo es más caliente que la piel, el cuerpo gana calor por convección y radiación, y pierde por evaporación al sudar.
La radiación solar absorbida por el cuerpo humano es bastante variable y pequeña en relación a la generación interna de calor, sin embargo en un día soleado y sin viento, puede llegar a ser significativa.
Variables como: declinación solar, turbidez atmosféricas, cubierta nubosa, latitud, reflectividad ambiental, área de exposición, reflectividad y absorbitividad de la ropa, afectan la entrada de energía al cuerpo.
Una persona en reposo, en promedio inhala aproximadamente 250 ml de oxígeno por minuto y exhala la misma cantidad de CO2 .
El metabolismo basal humano crea calor a una razón de 2000 KCal/día, la cual puede duplicarse cuando se realizan labores manuales pesadas.
El balance de agua y calor nos indican las respuestas del organismo, sin embargo un conocimiento de las magnitudes de sus fuentes y sumideros de energía será necesario, por lo que la energía del proceso metabólico puede ser dado por:
Qm = QB + Qp = Q* + QH + QE
Donde QB es el metabolismo basal y Qp la producción muscular de calor.
Un Clo se define como el aislamiento térmico por la ropa que mantiene el confort de una persona en reposo dentro de la casa, con temperatura del aire de 21°C y humedad relativa menor de 50°.
La "temperatura efectiva" se define como el índice en que la temperatura (sin efectos de ventilación, con aire saturado) induce en una persona sedentaria (con ropa ordinaria dentro de la casa), la misma sensación de confort como la inducida por las condiciones reales del momento de temperatura, humedad y movimiento del aire.
POTENCIALES CLIMÁTICOS DE PRODUCCIÓN PRIMARIA Y ESTABILIDAD PRODUCTIVA
La riqueza de un país está constituida, en parte, por sus potenciales de productividad primaria, es decir, por la propiedad natural que posee la vegetación de captar la energía solar incidente y mediante la fotosíntesis, generar substancias orgánicas susceptibles de ser utilizadas como materiales nutritivos.
La productividad primaria de un ecosistema se refiere a la velocidad a la que se almacena la energía como resultado de la actividad fotosintética o quimiosintética de organismos productores (principalmente las plantas verdes) en forma de substancias orgánicas (Odum, 1971).
Es una medida de acumulación de energía en un período específico y en una área determinada (Jones, 1979).
La constante conversión de energía luminosa en energía química es de gran importancia, ya que la mayor parte de los organismos no fotosintetizadores, como el hombre, dependen de los autótrofos para obtener la energía que los mantiene vivos.
De esta manera el crecimiento de la vegetación, además de tener importancia biológica adquiere implicaciones económicas y políticas e influye en el desarrollo social.
Los potenciales de producción primaria en los ecosistemas terrestres son el inicio de una cadena de transferencias energéticas que desembocan en la disponibilidad de energía en forma de alimento, (Cooper, 1975).
Es lo que determina que en los ecosistemas se presenten diferentes grados de rapidez en el almacenamiento de esa energía, y tanto los potenciales de producción, como la capacidad de respuesta y eficiencia, son una resultante de perturbaciones ambientales estudiadas por la microclimatología, (Lowry, 1970).
Los potenciales de productividad primaria varían de una región a otra, encontrándose áreas de baja productividad, como los desiertos, o zonas de alta productividad como las selvas.
Las causas de estas diferencias residen en las características genéticas, fisiológicas y bioquímicas de las plantas, así como en la relación de éstas con otros organismos y su medio ambiente, en donde las interacciones y combinaciones posibles de factores como la radiación solar y sus derivados (luz y temperatura), precipitación y tipo de suelo, ejercen una notable influencia sobre los patrones de distribución y crecimiento de la vegetación.
Los estudios de la radiación neta total en superficies naturales, es decir, sobre el suelo descubierto, los pastizales, los cultivos y los bosques, resultan importantes, ya que representan la cantidad de energía disponible para los procesos de fotosíntesis y transferencia de calor latente y sensible, como componentes del balance de calor entre estas superficies y la atmósfera (Billings, 1978).
Estas estimaciones se utilizan también para el cálculo de la evaporación en estudios hidrológicos y de balances de energía en estudios climatológicos.
Son muchos los factores que pueden reducir o incrementar la productividad, tanto en los ecosistemas naturales, como en los cultivados; entre estos factores se consideran importantes la radiación solar, el dióxido de carbono, la temperatura, el agua, el nitrógeno, el fósforo y pequeñas cantidades de ciertos minerales, etc. (Gates, 1971).
También son importantes las fuentes externas, que aportan como subsidio energía que se origina fuera del sistema, lo que incluso reduce los costos de mantenimiento.
Estas fuentes externas que subsidian energía pueden estar representadas por el viento, la lluvia, la energía contenida en un combustible fósil o en el trabajo animal o humano (Margatef, 1981).
Sin embargo, un mismo factor que en determinadas condiciones actúa como subsidio, reforzando la productividad, en otras puede generar pérdidas, tal como sucede en la insolación, vientos, precipitación, etc. (Odum, 1971).
Los estudios microlimáticos son necesarios para conocer los mecanismos de adquisición, almacenamiento y disponibilidad de energía, tanto en los sistemas naturales como en los agrosistemas manejados por el hombre,(Rosenberg, 1974).
Así de esta forma, podemos conocer cuánta energía nos proporciona la naturaleza y con qué rapidez nos la cede; al igual que las fluctuaciones y sus valores promedio, sus mínimos y máximos y lo que es mejor, su óptimo, importante y necesario para el manejo de cualquier recurso natural en explotación, (Munn.1970).
Con el propósito de evaluar la productividad primaria en los sistemas ecológicos, se han diseñado algunos métodos de medición, la mayoría usados en alguna cantidad indirecta, como por ejemplo, el método de la cosecha, en el que se recoge y pesa la cantidad de materia vegetal producida durante un determinado período (Kira et al., 1967; Odum, 1971; Billings, 1978, Wielgolaski, 1975).
Otro método consiste en medir las velocidades de fotosíntesis y respiración de plantas completas o áreas con vegetación, utilizando algún instrumento que mida la absorción o liberación de dióxido de carbono u oxígeno en cierto tiempo (Egle y Sohenk, 1951; Billings et al., 1966).
Se pueden utilizar también métodos fundados en la circulación de átomos radiactivos o en la cantidad de clorofila por metro cuadrado (Billings, 1978).
Estos y otros métodos se apoyan en principios relativamente sencillos, pero su aplicación práctica es frecuentemente muy compleja para poder obtener resultados exactos, especialmente cuando se aplican a ciertos tipos de vegetación, como bosques, selvas, etc. (Jones, 1979).
En este tipo de métodos se consideran los aspectos relativos a la estructura y características anatómicas y fisiológicas de las plantas.
Algunas propiedades ambientales también se destacan por su importancia y entre ellas procede mencionar: las radiaciones solar y térmica, la temperatura del aire, la humedad relativa, la precipitación, el viento y el balance de radiación (Helms, 1965; Gates, 1965; Botkin, 1969; Miller, 1970, Vocum et al., 1964; Monteith, 1965; Budyko, 1974; Sneva, 1982).
De acuerdo con un enfoque diferente, algunos investigadores se inclinan por la construcción de modelos donde se consideren tanto la contribución de algunas variables ambientales, como la influencia de las características de la vegetación, en cuanto a la productividad.
La influencia de cualquier factor ambiental en el campo de la producción agrícola, no puede considerarse en forma aislada.
Sin embargo la radiación recibe una atención especial, ya que sus balances dan la disponibilidad de energía para el desarrollo del microclima de las plantas e influye directamente en las respuestas fisiológicas de los vegetales, (Oke, 1978)
La radiación tiene un efecto directo sobre la convección, evaporación y fotosíntesis; e indirecto en el control de la apertura estomática junto con otros factores, tales como la extensión del mezclado vertical, la velocidad del viento y el abastecimiento de agua al suelo, y es responsable de la distribución vertical de la temperatura, de la humedad del follaje y de las temperaturas, tanto del suelo como del vegetal (Budyko,1974).
Los cultivos tienden a crear su propio microclima como respuestas a estas interacciones lo que provoca diferentes grados de "stress", resultante en una mayor o menor producción.
Es necesario en el desarrollo de la agricultura, seleccionar las áreas en donde las características climáticas sean adecuadas, a fin de obtener de cada cultivo, índices de producción óptima.
Al respecto, se han llevado a cabo diversas investigaciones que señalan la influencia directa de la variación de los parámetros meteorológicos en la producción agrícola, (Michalczy, 1979).
Se ha estimado que la biomasa del fríjol de campo (Vicia faba L.), está relacionada con la cantidad de radiación fotosintéticamente activa absorbida y con la humedad del suelo (Fasheun, .1982).
Para México, (Ritter et al 1986) determinaron los potenciales de producción primaria en diferentes ecosistemas naturales, en función de la disponibilidad de agua y energía en cada sistema y la estabilidad como un potencial energético latitudinal.
MODELOS CLIMÁTICOS DE PRODUCTIVIDAD PRIMARIA
Con apoyo en la idea de que el ambiente determina en cierta forma las características de las comunidades terrestres, algunos autores sostienen que es posible establecer correlaciones entre valores de variables ambientales y cifras de productividad.
Los registros que se obtienen de las estaciones meteorológicas constituyen una importante fuente de información que se puede utilizar para tratar el problema, basándose en el criterio de las posibles asociaciones entre la variación ambiental y la productividad.
(Rosenzweig 1968) ha señalado que la evapotranspiración real (ER), es decir, la cantidad de agua evaporada del suelo y transpirada por la vegetación , se puede utilizar en la predicción de la productividad primaria neta.
Esto deriva de que la evapotranspiración real expresa simultáneamente la disponibilidad de agua y de energía solar, ambos, elementos indispensables para la fotosíntesis.
Este autor utiliza un conjunto de datos de productividad primaria neta (únicamente sobre el suelo), correspondientes a diferentes tipos de vegetación (bosque, desierto, pastizal, tundra y bosque tropical).
Estos datos los relaciona con los valores de evapotranspiración real, calculados por el método de Thornthwaite y Mather (Thornthwaite y Mather, 1957), citados por ( Rosenzweig,"1968).
El modelo de Rosenzweig se apoya en un análisis de regresión lineal, comprendiendo en los términos de la ecuación a los logaritmos de las variables.
PER1 = log NAAP = (1.66 + 0.27) log ER - (1.66 + 0.07)
Donde:
ER = evapotranspiración real en mm/año
NAAP = productividad primaria neta sobre el suelo g/m2/año.
Rosenzweig considera que la velocidad de fotosíntesis depende de las concentraciones de materia prima comprendiendo entre otras el agua y la radiación solar.
En los ambientes terrestres, el CO2 es más o menos constante (Sellers, 1965), citado por (Rosenzweig, 1968).
Así la evapotranspiración real, según el autor, es una medida de los elementos más variables.
Se afirma que existen condiciones locales especiales que ningún modelo general toma en consideración, tales como la deficiencia de algún nutrimento esencial, como el fósforo o el nitrógeno, o condiciones locales de neblina que tienden a elevar la concentración de CO2 y por lo tanto la productividad (Wilson, 1948), citado por (Rosenzweig, 1968).
La medida restringida sólo a la evapotranspiración real, no podría abarcar estas variables para la determinación de la productividad (Lieth y Box, 1972) en general, siguen la misma lógica que sirve de fundamento a Rosenzweig, como él, consideran a la productividad primaria como función de la evapotranspiración real, sin embargo, existen diferencias en cuanto al conjunto de datos, ya que para la elaboración de su modelo utilizan valores de productividad primaria neta total (incluyen datos de superficie y por debajo del suelo) en América, Eurasia y África, combinados con valores del mapa de Geiger de evapotranspiración y con apoyo en los mismos, construyen la ecuación.
PER2 = 3 000 (1 - e-0-0009695 (ER-20)
Donde
PER2 = Productividad primaria neta en g/m2 /año
Con este modelo, llamado "Modelo Montreal 2, conmemorativo de C.
W. Thornthwaite", se calcula que el valor global de la productividad primaria neta trimestre es de 118. x 109 ton/año.
(Lieth, 1973), por su parte, destaca la importancia de la temperatura media anual, T oC) y de la precipitación media anual, Pr (mm), como factores ambientales útiles para la determinación de la productividad (PT, PN).
Los parámetros considerados se combinan para la elaboración del "Modelo de Miami", que se expresa en dos ecuaciones:
PT =
Para la temperatura, frente a la productividad (g/m2 /año) y
PN - 3 000(1 - e0-000664Pγ)
Para la precipitación frente a la productividad en la base de los logaritmos naturales.
Las ecuaciones obtenidas se basan en un conjunto de lecturas procedentes de más de mil estaciones distribuidas sobre los continentes, que comprenden información sobre temperatura y precipitación; los datos se transforman en valores PPN, seleccionando el valor más bajo cuando las dos ecuaciones producen resultados diferentes.
DETERMINACION DE LAS RELACIONES CLIMA-PRODUCCION.- Son muchos los factores que pueden reducir o incrementar la producción tanto en los sistemas naturales como en los cultivados, como son la radiación solar, el dióxido de carbono, la temperatura, el agua y los nutrimentos).
Esta, para los sistemas naturales varía de una región a otra, encontrándose áreas de baja producción como los desiertos o zonas de alta producción como son las selvas.
Las diferencias son debidas a características biológicas intrínsecas de los vegetales y a su microclimatología de desarrollo.
Bajo esta última idea se propone la siguiente consideración: Debido a que el ambiente determina en cierta forma las características de los ecosistemas terrestres podemos desarrollar modelos climáticos de producción vegetal y establecer relaciones de las variables climáticas con valores de producción.
Entre las variables climáticas a relacionar con la producción, el índice de aridez de (Budyko, 1974), tiene la conveniencia de unificar en un sólo valor la disponibilidad energética y de agua, factores determinantes en la producción de cualquier cultivo o ecosistema.
MODELOS MATEMÁTICOS DE PRODUCCIÓN VEGETAL.- Las siguientes consideraciones físicas son deseables en los modelos de simulación matemática de producción vegetal:
- Una descripción matemática en forma tridimensional de dos campos de radiación, CO2, agua, temperatura, viento, etc.
- Parametrización de los procesos de intercambio de masa y energía entre las plantas y su micro ambiente.
- Establecer relaciones de la dependencia de fotosíntesis y respiración de factores físicos, químicos y biológicos.
- Descripción matemática de la distribución y orientación de los elementos del follaje de la planta (altura, índice de área foliar, distribución vertical del área y la masa de los diferentes órganos de la planta, su función de distribución de área foliar en términos de su inclinación y orientación y la dispersión horizontal del área foliar.
CALCULO DE LA ESTABILIDAD PRODUCTIVA.- Para determinar la estabilidad productiva de los diferentes sistemas de estudio, se tomaron en cuenta las consideraciones establecidas por (May, 1973), las cuales en el caso de una simulación matemática determinista son: Para pequeñas fluctuaciones en la población, la dinámica se describe por la matriz comunitaria y el tiempo de retorno al equilibrio es medida por lo negativo de las partes reales de su eigenvalor (λo).
La dinámica de las interacción biológicas (λ = ΣReal (λo) max) tienden a restaurar a la población al punto de equilibrio, mientras que las fluctuaciones del ambiente actúan para alejarla.
En un ambiente con fluctuaciones climáticas aleatorias (σn2), la estabilidad biológica es dada por λ mayor que 0, pero si λ es mucho menor que la varianza (σn2) la población sufrirá grandes fluctuaciones, dirigiéndola a su extinción.
Pero si λ es mucho mayor que σ
n2, las fluctuaciones poblacionales observadas son relativamente pequeñas.Por lo que la estabilidad o la inestabilidad de la población depende del balance del poder entre la estabilización biológica y la aleatoriedad de las fluctuaciones ambientales, (Ritter et a1. 1988).
COMPORTAMIENTO, USOS Y APLICACIONES MICROCLIMATICAS
EN LA PRODUCCIÓN AGRÍCOLA.
El hecho de que se esté iniciando una serie de años de sequía en zonas que son los graneros mundiales hizo que los principales climatólogos se reunieran en Bonn, Alemania en 1974.
La conclusión a la que llegaron es que efectivamente la tierra está experimentando cambios climáticos que pueden ser desastrosos para la producción agrícola.
La oleada actual de catastróficas sequías va ligada a las expansiones de lo que se conoce como los "Vórtices circumpolares" (vientos de origen polar de gran altitud) que pueden afectar a la climatología casi hasta el Ecuador.
Si no nos anticipamos a estos cambios climáticos, haciendo un mejor uso de nuestros granos, así como incorporando las técnicas para un mejor conocimiento y uso del Microclima en que se desarrollan los cu1tivos, existe la posibilidad de que podrían morir de hambre hasta mil millones de personas en los años venideros.
Esto, aunado a que el hombre cultiva hasta la última hectárea que pueda ser provechosamente utilizada de este mundo, y de que sólo un 7% de esta masa de tierra cuenta con la adecuada combinación de lluvia, temperatura y elementos nutritivos que permitan el tipo de agricultura Occidental, ya que la mayor parte de la tierra cultivable no se presta a ser mecanizada; de que 460 millones de gentes sufren deficiencia nutricional y de que la población aumenta entre 75 y 90 millones anualmente, necesitándose 280 millones de hectáreas de nuevas tierras para alimentar a esta población que nace.(Pirie,"1969).
Así también el exceso de laboreo y pastoreo ha estropeado más tierras de las que el hombre ha añadido a su reserva durante el último siglo.
La irrigación y los desmontes han creado más desiertos que nuevas tierras de cultivo, se ha provocado una salinización de los campos y un deterioro irreversible, o muy costoso para ser operativo, y millones de hectáreas han tenido que ser abandonadas.
Esto aunado a que gran parte de la tierra de los trópicos son suelos lixiviados que no merecen la pena de ser cultivados.
La esperanza de los cereales híbridos producidos con la "Revolución Verde" son también una desilusión, ya que éstos precisan de gran cantidad de agua y fertilizantes y no son resistentes a las plagas y producen menor cantidad de proteína,(Brill, 1981).
De todo esto se puede concluir que la naturaleza tiene límites y que el hombre ha perdido la capacidad de prever y prevenir y que finalmente terminará destruyendo la tierra.
La única esperanza se encuentra en la modificación radical de nuestra conducta, pero tal parece que el hombre cuando se enfrenta a una situación real desagradable en lugar de afrontar y tomar una solución adecuada que podría representar el sacrificio de nuestro bienestar personal, simplemente lo apartamos del pensamiento y actuamos como si tal contratiempo no existiera.
La destrucción de la naturaleza es la mayor amenaza para la sobrevivencia y bienestar de la población y rara vez estamos conscientes de esta realidad, pero obviamente el fracaso de la conservación de los recursos vitales se encuentra íntimamente relacionado con el surgimiento de otros problemas.
Si no se pone el énfasis necesario en la conservación de la naturaleza, la vida resultará progresivamente más difícil y completamente imposible a los marginados.
Cuidar la fertilidad del territorio nacional y la calidad de nuestros satisfactores es indispensable ya que el no hacerlo es una de las formas más funestas de corrupción, aunque la causa sea la ignorancia y no el "lucro, (Cesarman,:1972).
A nuestra falta de responsabilidad, se va agregando el hecho de que nuestras mayores áreas de .producción agrícola en la República Mexicana, son de temporal, lo que nos da una riesgosa dependencia en la fragilidad productiva de estos sistemas, que están en función de la aleatoriedad climática, el cual se espera sufra severas modificaciones y en cuya aceleración de presencia no estamos exonerados de culpa.
Debemos prever resultados anticipándonos a las posibles respuestas de la naturaleza y de su magnitud de reacción, debemos conocer cómo el microclima influye en algo tan básico y esencial como es la producción agrícola, de cómo usarlo para aumentar la eficiencia productiva de las plantas, de sus posibles modificaciones, creando ambientes con un mayor grado de confort, permitiendo así una mayor energía a funciones naturales de producción y una mínima a las de sobrevivencia.
A la agricultura la podemos definir como superficies grandes y poco rentables para la recolección de la luz solar, existiendo otros sistemas más eficientes en la producción como el nutricultivo que consiste en un cultivo en una solución acuosa, con rendimientos de 6 a 8 veces superior al cultivo tradicional, así también las algas del género chrorella se pueden cultivar en grandes tanques y son ricas en proteínas, grasa y vitaminas, además parecen tener todos los aminoácidos necesarios con rendimientos de 40 veces la planta más proteínica (la soya) y .160 veces la carne de ternera.
De todo esto la única solución posible frente al siglo XXI es la irrupción de nuevas técnicas capaces de ser utilizadas en gran escala.
Las materias primas básicas para la industria del futuro serán: el agua del mar, el aire, las rocas comunes, los depósitos sedimentarios de rocas calizas y fosfatadas y la luz solar.
Todos los ingredientes esenciales para una sociedad altamente industrializada se encuentran presentes en la combinación de estas sustancias, (Kahn, H.y Weiner, 1967).
Del agua del mar se obtendrá el agua para la agricultura y la industria, hidrógeno para la reducción del mineral de hierro, cloro, sosa cáustica, magnesio, sal, bromo, yodo y potasio, junto con pequeñas cantidades suplementarias de otros elementos.
Del aire se extraerán ácido nítrico y amoníaco, que serán utilizados para la producción de comida y en procesos industriales, Oxígeno y otros gases necesarios a la industria también provendrán del aire.
De la roca común se obtendrán la mayor parte de los metales esenciales y las materias fisionables.
De la roca caliza se extraerá el carbón, importante punto de partida para la industria de la química orgánica y para la fabricación de carburantes líquidos.
De las rocas fosfatadas se obtendrán fertilizantes.
De la luz solar, electricidad y una amplia variedad de sustancias orgánicas que se utilizarán como alimento y como elementos esenciales para varias industrias químicas.
INFLUENCIA DE LA RADIACIÓN SOLAR.- La cantidad de radiación solar que reciben las plantas es de gran importancia para la producción agrícola.
Las plantas dependen vitalmente de la radiación solar y gran parte de la energía recibida la utilizan para su transpiración y crecimiento.
La energía solar ejerce considerable influencia sobre la composición química de las plantas y sus frutos.
El contenido de azúcar en la remolacha, por ejemplo, depende del número de días con sol que haya estado expuesta durante el período de crecimiento, y cuanto mayor sea el número de días con sol mayor será el contenido de azúcar.
La misma sensibilidad al número de días con sol es típica de la uva, pues su contenido de azúcar y otras cualidades así como la calidad del vino elaborado, dependen de la cantidad de energía solar recibida por las plantas durante el período de crecimiento, y mientras más energía reciba, mejor será la calidad.
Cuando la radiación solar es insuficiente se puede producir una flexibilidad deficiente de los tallos de las plantas, observándose en los cereales que no son capaces de erguirse por si mismos después de fuertes vientos.
La radiación solar puede ejercer algunas veces una influencia perjudicial, como en el caso de la papa, la que, después de la exposición de la luz, se tornan verdes, acumulan un alcaloide (la solanina) y se convierten en no aptos para el consumo humano, aunque la calidad, como semilla, se conserva y hasta mejora.
La exposición de los tubérculos a la radiación solar aumenta su resistencia de conservación durante el almacenamiento, (Vitkevich, 1960).
La radiación solar puede también llegar a retardar el crecimiento de las plantas, observándose un crecimiento más rápido durante la noche y en las zonas montañosas, las plantas llegan a desarrollar órganos especiales de protección contra las fuertes radiaciones, evitando así una evaporación excesiva.
Las plantas que crecen en la sombra tienden a desarrollarse más en sentido horizontal, pero en general las plantas tienden a tomar direcciones diferentes de acuerdo a las condiciones de iluminación, escogiendo una dirección apropiada con el propósito de recibir la mayor cantidad posible de luz.
Sin embargo en las regiones áridas las plantas tienden a presentar al sol los bordes de las hojas recibiendo así solamente una pequeña porción de la energía solar por unidad de área, para reducir al máximo sus pérdidas de agua por evaporación, (Cooper, 1975).
En países montañosos como el nuestro, donde la topografía influye en el régimen solar y en el de los otros elementos microclimáticos, significa que los campos de cultivo por diferentes orientaciones y pendientes tienen horas diferentes de insolación y reciben cantidades distintas de radiación solar.
El exceso de la radiación incidente, característica en gran parte de nuestro país, hace que consideremos sobre cómo hacer un mejor uso en la agricultura de este recurso.
Una técnica prometedora puede resultar la siembra de diferentes cultivos con una estratificación en las alturas que nos permita una mayor absorción de insolación en todos los niveles posibles.
REGULACIÓN DE LA RADIACIÓN SOLAR RECIBIDA POR LAS PLANTAS Y UTILIZACIÓN TÉCNICA DE ESTA ENERGÍA.- En la mayoría de las zonas áridas, el problema radica mas en reducir que en incrementar la radiación.
Para muchas plantas, la cantidad de energía solar recibida bajo condiciones de campo puede aumentar o disminuirse hasta cierto punto, si se modifica la dirección de la plantación o siembra, se ha demostrado que esto afecta profundamente la cantidad y calidad del rendimiento de los cultivos.
Un incremento de ésta puede efectuarse mediante la reducción del número de semillas sembradas por unidad de área, o también mediante el aclareo de plantas o podar en el follaje de los frutales, teniendo el cuidado de plantarlos lo suficientemente alejado uno del otro.
Se puede obtener una reducción en la cantidad de energía recibida por cada planta individualmente al aumentar la densidad de la siembra o la norma de siembra, o por la siembra de plantas de tallo largo en filas protectoras, o por el cultivo de plantas bajo la sombra de árboles y por otros métodos.
Debemos de considerar que las plantas no solamente requieren de energía solar, sino de un conjunto complejo de condiciones y particularmente, un área determinada de alimentación.
La ventaja de una dirección Norte-Sur disminuye cuando nos alejamos del Ecuador, lo que se debe a que, con el aumento de la latitud, el Sol ocupa una porción cada vez mayor del arco del horizonte, elevándose y poniéndose cada vez más cerca del Norte.
Así, cuando más nos acercamos hacia el Sur, mayor es el contraste entre las hileras de siembra Norte-Sur, y Este-Oeste, en relación con la cantidad de radiación solar recibida por las plantas.
Otro punto importante es el siguiente: Las plantas que se encuentran en hileras Este-Oeste están afectadas por los rayos del medio día, esto es, por una concentración alta de ondas cortas violeta y ultravioleta y las plantas situadas en hileras Norte-Sur están afectadas fundamentalmente por los rayos de la mañana y de la tarde, o sea, por una concentración menor de ondas cortas.
En otras palabras, las hileras de plantas orientadas en dirección distinta no solamente difieren en la cantidad de radiación solar recibida, sino también en la composición espectral de esta radiación: (Stanhill et al. ,1965).
Así pues, se llega a las conclusiones siguientes:
1. Los surcos e hileras que tienen dirección Norte-Sur reciben considerablemente más calorías que los que tienen dirección Este-Oeste, a través de toda la estación del crecimiento y en todas las latitudes.
2. La diferencia en la cantidad de calorías recibidas aumenta hacia Junio, o sea, hacia el solsticio de verano, durante el cual los rayos del sol tienden a la verticalidad máxima.
3. La diferencia de las calorías recibidas entre las dos direcciones de siembra es mayor en las regiones del Sur, o sea, en latitudes bajas.
4.Una hilera de plantas con orientación Este-Oeste no tendrá la misma cantidad y calidad en el rendimiento agrícola, que una hilera con orientación Norte-Sur.
5.La producción obtenida en hileras Norte-Sur, es mayor o menor y de buena o mala calidad, de acuerdo con el cultivo dado y sus necesidades de radiación. Esta conclusión puede aplicarse a hileras simples, pues en los campos, desde luego, cada hilera está parcialmente sombreada por su vecina y la influencia de la dirección es algo menor. No obstante, la dirección Norte Sur mantiene su superioridad.
TEMPERATURAS DEL AIRE Y DEL SUELO.- Las temperaturas del aire y del suelo son los principales factores que controlan la tasa de las reacciones químicas que implican la fotosíntesis y el crecimiento de las plantas.
Todas las plantas de cultivo requieren que el suelo tenga una cantidad determinada de calor, si han de producir un rendimiento óptimo.
La influencia de la temperatura del suelo sobre las plantas es de importancia considerable, principalmente durante la etapa de crecimiento.
Para que las semillas germinen es necesario que la temperatura del suelo permanezca dentro de ciertos límites que "correspondan a los requerimientos de la planta dada".
La duración en el crecimiento y desarrollo de un cultivo, se observa que no es constante, variando según las regiones y hasta en una misma loca1idad, de acuerdo a los años y fechas de siembra.
En algunas variedades de trigo, se verá que desde la germinación hasta la maduración transcurren 142 días, en otras 155 días, 177 días, etc.
Si desde el momento en que se produce la germinación se suma la temperatura media de cada día hasta el momento de la madurez, la suma total es siempre la misma, cualquiera que sea la ubicación del cultivo y el año considerado.
La cebada requiere aproximadamente una suma de 1700°C, el trigo 2000°C y 2500°C el maíz.
La temperatura mínima del suelo a la cual el girasol puede germinar es de 5°C, aunque las temperaturas mayores son más favorables y tienden a acelerar la germinación (la temperatura óptima es la comprendida entre 31 y 37°C); después de 37°C comienza a disminuir el porcentaje de germinación y los aspectos del vigor germinativo, (Vitkevich, 1960).
La temperatura del suelo es especialmente crítica cuando éste está muy húmedo o cubierto con una lámina de agua, ya que el agua obstruye la transferencia entre el aire del suelo y la atmósfera exterior, lo que produce una deficiencia en oxígeno que conduce al amarillamiento y muerte de las plantas.
La temperatura del suelo también tiene influencia considerable en el desarrollo del sistema radical.
En el caso de los cultivos de invierno, por ejemplo, el desarrollo adecuado de las raíces tiene lugar entre los 6 a 10°C.
Las disminuciones graduales de las temperaturas del suelo y el aire, hacen posible que las plantas sinteticen el azúcar y se desarrollen con un vigor considerable. En cambio, las disminuciones bruscas de temperatura disminuyen el vigor de las plantas.
La papa cultivada a una altitud de 3,800 metros, puede soportar temperaturas tan bajas como 7. y 8°C bajo cero, pero en las planicies muere a 2°C bajo cero, esto se debe a que las plantas que están bajo la influencia de radiación solar intensa, que ocurre en las zonas de gran altitud, elaboran mayor .cantidad de azúcar.
En suelos húmedos y porosos las raíces necesitan poca energía para su desarrollo, y poco oxígeno para su óptimo crecimiento, ya que la compactación del suelo restringe el crecimiento de las raíces y de la planta.
Las temperaturas del suelo intervienen en las actividades fisiológicas y en la morfología de las raíces, siendo las temperaturas altas las que promueven la floración y acortan la etapa vegetativa, alcanzando las raíces un crecimiento óptimo cuando la temperatura está alrededor de los 20°C, obteniendo con ésta además los máximos valores de absorción de nutrientes.
La estructura del suelo afecta a la planta a través de la aereación, compactación, agua y temperatura, observándose en zonas desérticas que la nula precipitación produce poco intemperismo químico y poca formación de arcilla y materia orgánica, así como también poca vida microbiana.
Donde la evaporación y la precipitación tienden a igualarse existen buenas condiciones para la formación de arcillas y microorganismos.
En climas húmedos y fríos permiten también una abundante formación de arcillas y crecimiento vegetal, pero las arcillas y los nutrientes no se pueden acumular por lixiviación excesiva.
Las mayores lixiviaciones coinciden con las altas temperaturas donde el agua de drenaje se vuelve alcalina por rápida descomposición de la materia orgánica.
El contenido de materia orgánica tiene una gran influencia sobre la lixiviación, ya que aumenta la capacidad de retención de humedad.
Los suelos de textura gruesa tienen baja capacidad de retención de agua y los de textura fina lo contrario.
Así los cambios climáticos pueden ser también importantes dentro de la historia de los suelos, los cuales cambian en respuesta a los cambios en su ambiente.
INFLUENCIA DE LA CUBIERTA VEGETATIVA EN EL RÉGIMEN TÉRMICO DEL SUELO.
La cubierta vegetal tiene una influencia considerable en el régimen térmico del suelo, observándose que la temperatura en un suelo cubierto con vegetación difiere apreciablemente del de uno sin vegetación.
La presencia de vegetación afecta el equilibrio térmico del suelo y del aire superficial de una manera muy compleja.
La cubierta vegetativa no solamente reduce la afluencia de energía solar a la superficie del suelo, sino que también reduce la pérdida de calor por emisión durante las horas diurnas y especialmente durante la noche.
Las mediciones de temperatura del suelo realizadas a varias profundidades en terrenos cultivados y sin cultivar, indicaron que el calentamiento y la penetración de calor es más profundo en los suelos sin cultivar que en los suelos cultivados.
La diferencia de temperatura entre los dos depende de las propiedades del suelo, la profundidad, el tiempo y los climas locales.
En la transpiración de las plantas se absorbe parte de la energía solar que de otra manera produciría calentamiento del aire.
El resultado es que las temperaturas del aire, tanto dentro como encima de la cubierta vegetal son más bajas que las que se observan sobre las superficies del suelo desnudo bajo condiciones meteorológicas similares.
Las más altas temperaturas del aire dentro del follaje, se encuentran generalmente al nivel donde ocurre la máxima absorción de la radiación, (Lowry, 1970).
En zonas muy densas, la radiación solar jamás alcanza el suelo, éste recibe solamente la radiación reflejada y emitida por las hojas y la que penetra a través de ellas.
Las conclusiones que pueden hacerse son, que las plantas tienen una gran influencia en la temperatura del suelo, observándose que el terreno de un campo boscoso tiene durante el verano una temperatura inferior y en el invierno superior a la temperatura de un terreno sin cubierta forestal.
Correspondientemente, la temperatura media anual de la superficie del suelo y la amplitud de las oscilaciones anuales, son menores en los terrenos con bosques.
HUMEDAD DEL AIRE Y SUELO.- En el proceso de crecimiento de las plantas se requiere la presencia de agua en las células de la hoja, así como también la penetración dentro de la hoja del bióxido de carbono atmosférico.
Esta penetración es posible por los estomas de la hoja que se abren bajo el influjo de la luz del sol pero con la consecuencia de que el vapor de agua se escapa hacia la atmósfera.
La fuente normal de agua para la vegetación es el suelo, de donde la extraen las plantas por medio de sus sistemas de raíces.
En las regiones áridas, algunas plantas parecen ser capaces de utilizar el rocío.
Debemos considerar que la humedad es el principal factor limitante para la productividad agrícola en nuestro país por lo que un buen conocimiento del comportamiento de sus elementos siempre es deseable, como lo sería su conservación a fin de prolongar su presencia que nos permita asegurar un mayor número de cosechas durante el año.
Las plantas durante el día a través de sus estomas, sufren de una rápida pérdida de agua por transpiración.
El primer efecto de la cubierta vegetal es reducir la evaporación directa desde la superficie del suelo por la sombra y por la restricción del movimiento del aire.
Para compensar esto las plantas tienen que ser capaces de extraer agua del suelo a una velocidad superior a la que normalmente ésta se difunde.
La rapidez de pérdida de humedad del suelo está muy influenciada por la cantidad y tipo de vegetación existente.
La "Filtración" del flujo de radiación solar en la vegetación tiene dos efectos directos sobre las plantas en los niveles más bajos.
El primero es reducir la tasa de la fotosíntesis, el segundo reducir la de la transpiración, independientemente de la temperatura del aire, (Budyko, 1974).
Las hojas de cacao a la intemperie transpiran aproximadamente más que las plantas similares que crecen en un 90% de sombra.
Las condiciones favorables para el crecimiento del cacao se encuentran entre un 25% un 50% de sombra, ya que la intensidad de la fotosíntesis se reduce cuando la sombra excede del 50%.
El agua tiene una capacidad térmica específica elevada (un cal g-1 °C), mientras que el suelo seco tiene una capacidad térmica específica próxima a 0.2 cal g-1 °C.
En las mismas condiciones climáticas, un suelo arenoso que contenga solo 15% de agua a la capacidad de humedad de campo, se enfría y calienta con mayor rapidez que un suelo arcilloso que retenga 30% de agua.
La evaporación es mínima en suelos cuyas partículas tienen diámetros de 0.5 a 5mm, en suelos más finos el movimiento capilar del agua es mayor, mientras que en suelos gruesos, el aire tiene más libre acceso a las superficies de evaporación.
El incremento de la transferencia de agua del suelo a la atmósfera debido a la vegetación significa que una mayor proporción del flujo de radiación recibido se utiliza en la evaporación y una parte menor para el calentamiento del suelo, la vegetación y el aire superficial.
La vegetación tiende a crear sus propias condiciones favorables, ya que la transpiración de una planta rodeada de otras será menor que la de una que crece sola.
La presencia de las plantas modifica la estructura del suelo y reduce el escurrimiento superficial con el resultado de que una parte de la precipitación que alcanza el suelo se infiltra en él.
Por otra parte, la vegetación puede, en ciertas circunstancias incrementar la precipitación.
Esto sucede en áreas donde las nieblas bajas son frecuentes o sobre montañas suficientemente altas para estar frecuentemente entre las nubes.
Particularmente si hay viento, la niebla o las gotas de las nubes son recogidas por las hojas y ramas y alcanzan el suelo, ya sea por goteo o corriendo por los tallos, (Rosemberg,1974).
La proporción de la precipitación interceptada por la vegetación depende de su densidad, así como también de la "duración e intensidad de la lluvia o la nieve.
La vegetación reduce la cantidad de agua que alcanza a llegar al suelo por la intercepción de la lluvia, pero también juega un papel importante en la reducción de la erosión del suelo y el incremento de la infiltración.
La vegetación frena la caída de las gotas de lluvia y reduce así su efecto de erosión.
La vegetación actúa como obstáculo para el escurrimiento superficial en terrenos de declive y sus sistemas de raíces aumentan la permeabilidad del suelo.
Esto es particularmente importante en áreas montañosas donde las pendientes son empinadas, lo que puede hacer muy destructivo el escurrimiento superficial.
Existe otra forma en la cual las plantas pueden afectar el abastecimiento de agua al suelo y está relacionada con el rocío, ya que algunos experimentos de laboratorio parecen demostrar la absorción de humedad por medio de las hojas, cuando éstas se encuentran rodeadas de una atmósfera neblinosa o saturada.
La principal importancia de la vegetación en la vida de insectos y animales radica en su efecto moderador sobre el flujo de radiación, la temperatura del aire y en la protección que ofrece contra el viento y la lluvia (0ke, 1978)
Las amplias variaciones del microclima que pueden encontrarse dentro de la cubierta vegetal, permiten a los insectos y animales pequeños encontrar dentro de un corto radio de acción, las condiciones más apropiadas para su desarrollo.
MICROCLIMAS INTENCIONALMENTE MODIFICADOS.
MODIFICACIÓN DEL MOVIMIENTO.- Se puede Modificar la rugosidad de la superficie por medio de una vegetación alta o por estructuras artificiales.
En el primer caso se tiene la utilización de cortinas rompevientos y en el segundo la utilización de capelos o invernaderos.
Cortinas Rompevientos.- Las barreras de protección con árboles son usadas contra el daño físico a edificios y cultivos, pero la reducción en transferencia de calor puede también ser benéfica en conservar las pérdidas metabólicas de calor en el ganado y pueden reducir el consumo de energía en las granjas hasta de un 54%.
No menos significativo es el efecto combinado de las barreras que pueden incrementar el rendimiento en los cultivos.
Varios trabajos sugieren incrementos especialmente a distancias de 5 y 15 veces la altura, pero cerca de la barrera los efectos de sombra y competencia por humedad de los árboles de la barrera pueden disminuir el rendimiento para distancias de 0.5 veces su altura.
No todos los efectos de las barreras son benéficos por ejemplo, las bajas temperaturas nocturnas presentan un mayor riesgo de heladas y la combinación de vientos lentos y alta humedad pueden ayudar al desarrollo de enfermedades por hongos.
Respecto a la importancia de la permeabilidad de una barrera de vegetación para controlar el régimen del viento y su abatimiento, se debe a que una barrera muy densa es menos efectiva que una de densidad moderada, por la vigorosa turbulencia que ocurre detrás de la primera cuando el aire desciende en un 50% y puede fluir a través de ella, obteniéndose mejor protección contra esa turbulencia.
La velocidad del viento se reduce a menos del 80% de su valor en el campo abierto.
El área protegida por un rompevientos toma aproximadamente la forma de un triángulo obtuso cuyo ápice se mueve de acuerdo con la dirección del viento.
El espacio entre las cortinas debe medir generalmente de 25 a 30 veces la altura de los árboles.
La reducción de la velocidad del viento entre las cortinas de árboles afecta muchos aspectos del microclima, pero no hay duda de que ejercen un efecto favorable en el rendimiento de los cultivos, (Geiger 1969).
Debe tomarse en cuenta también qué tan serias pueden ser las condiciones que se ofrecen a la propagación de enfermedades de las plantas.
El aumento de depósitos de roció que se observa entre las cortinas de árboles puede proveer un ambiente favorable para el desarrollo de las enfermedades de las plantas.
Por otra parte, la protección que proporcionan dichas cortinas contra los vientos fuertes favorece la actividad de insectos polinizadores, mientras que su sombra beneficia a los animales domésticos, (Oke 1978).
Invernaderos.- Los beneficios técnicos de los invernaderos ha sido reconocido por mucho tiempo, sin embargo la explicación física permanece aún sin elucidarse.
Un invernadero presenta dos formas de control: primero, es un filtro a la radiación y segundo, reduce las pérdidas de calor por su protección contra el viento.
La propiedad del vidrio conocida como "Efecto de Invernadero", lo hace transparente a la radiación visible, pero opaca a los rayos infrarrojos, permite a la radiación solar entrar en el invernadero pero evita cualquier escape de radiación térmica a la Atmósfera.
El techo de vidrio intercepta, de hecho, una pequeña proporción del flujo de radiación arriba, correspondiente a la diferencia de temperatura entre sus superficies interior y exterior.
Esta diferencia de temperatura es una función del espesor del vidrio y su conductividad térmica.
Aún así, la radiación emitida por el techo debe ser mayor que la del terreno circundante, debido a la ausencia de evaporación a partir de la superficie del vidrio.
La protección proporcionada por los invernaderos es en realidad una protección contra el movimiento del aire.
El flujo de radiación no cambia mucho, pero la difusión turbulenta de calor y vapor de agua entre el suelo y la atmósfera está completamente suprimida, siendo reemplazada solamente por la conductividad térmica a través del techo y las paredes,(Lowry, 1970)
CONTROL DEL ALBEDO.- El albedo (definido como el cociente de la energía reflejada respecto a la incidente) es una propiedad superficial funda mental relativamente fácil de alterar.
Determina y regula directamente la capacidad de absorción del suelo.
Esto establece límites al balance de energía superficial y la interacción que tiene con el balance de agua, controlando la humedad y la temperatura del suelo y de la capa de aire adyacente al suelo.
El valor del albedo depende de factores como son la coloración, humedad
y rugosidad del suelo.
La manipulación del albedo implica una considerable reacción en cadena del clima y energías.
Coloración de Suelos.- La temperatura de los suelos ennegrecidos es más caliente que en los emblanquecidos.
En la noche el negro y el normal tienden a coincidir, pero no en el color blanco.
El suelo negro tendrá 50% menos humedad y el suelo blanco 50% más humedad que el normal.
Cambios de albedo inducen cambios que afectan como resultado su humedad, produciendo así cambios de evapotranspiración.
Control de la Geometría del Suelo.- El control de la geometría del suelo se usa en agricultura en la formación de "surcos" y "montículos"'.
En latitudes altas, la superficie nunca recibe la radiación solar directa en el cenit y así nunca reciben la máxima intensidad de radiación.
Con los surcos y montículos es posible recibir una mayor radiación, esto es especialmente útil en primavera cuando el calentamiento es crítico para el secado y germinado de las semillas cuando la elevación solar es baja.
La orientación del surco es también importante; en direcciones N-S ambas pendientes del montículo son irradiadas durante el curso del día, mientras que en orientaciones E-O, la parte sur es particularmente favorecida en el Hemisferio Norte.
Los surcos y montículos también actúan como trampas para aprisionar la luz y el calor.
Atrapando la radiación solar incidente, tiende a incrementarse la temperatura del suelo, y se tiende a reducir la superficie de enfriamiento, (Oke, 1978).
La influencia de la geometría de la radiación ha sido explotado en un amplio rango de aplicaciones prácticas; por ejemplo el espaciado de los surcos y la orientación puede manipularse para maximizar o minimizar la penetración de la luz y el atrapamiento de la luz y el calor.
Paredes o cercas de piedra coloreadas de blanco pueden ser usadas para emitir una mayor iluminación a las plantas y reducir su área de pérdida de radiación por la reflección y el calor.
La alta capacidad calorífica de las rocas transforman a las paredes en fuentes de calor en la noche.
Los colectores de energía solar para enfriar o calentar requieren de una orientación cuidadosa o aún, un medio continuo de seguir la trayectoria del Sol para optimizar su recepción, (Geigér, 1969).
La práctica de cubrir los suelos de paja es usualmente con el objeto de conservar la humedad del suelo al reducir la evaporación, pero puede también usarse para incrementar el calentamiento del suelo o prevenir el excesivo enfriamiento, dependiendo del tipo de cubierta usada.
La cubierta tradicional consiste de un material bien ventilado pero poco conductor.
Esto puede lograrse labrando la capa superior para introducir más aire o cubriendo la superficie con material aislante como el heno, paja, hojas, pedazos de madera, aserrín o grava.
Más recientes cubiertas artificiales han sido de uso común incluyendo el hule espuma, plásticos (opaco, translúcido o coloreado) papel y hojas de aluminio.
Como resultado de su bajo albedo, el plástico negro absorbe la radiación más eficientemente que el suelo desnudo y su temperatura sube hasta casi los 50°C.
Sin embargo, este calor no pasa fácilmente al suelo por el aire entre el plástico y el suelo que sirve de aislante.
La cubierta de plástico negro, entonces conserva la humedad y estabiliza la temperatura del suelo y actúa como una fuente de calor sensible para la Atmósfera.
Las superficies cubiertas de papel exhiben una disponibilidad de energía recibida en el suelo frente a ella y eleva la temperatura del aire y del suelo.
La modificación de la radiación y del equilibrio térmico durante el día puede lograrse con el uso de plantas más altas y mediante sombreado artificial.
La irrigación del suelo afecta el movimiento del aire, el calor específico, la conductividad térmica y el contenido de agua en las capas superficiales del suelo.
El flujo de calor hacia abajo durante el día en un suelo irrigado es más del doble de uno sin irrigación.
Lo que de hecho hace la vegetación, es distribuir la pérdida de calor a través de un volumen mayor de aire.
Se han empleado cortinas de humo para la protección de las huertas contra las heladas nocturnas, produciendo el humo de combustión incompleta del petróleo o por medios químicos.
Los principales inconvenientes del método del humo y que han conducido a su abandono virtual son sus efectos colaterales y el hecho de que, excepto en noches de perfecta calma, el humo es arrastrado por el viento y desviado del área escogida.
Para distribuir la pérdida nocturna de calor a través de una capa más profunda, de aire, se han empleado grandes ventiladores.
En noches tranquilas, el aire a unos metros encima de la vegetación es comúnmente varios grados más caliente que el aire a nivel del suelo.
Esta estructura térmica es muy estable, así que se necesita energía para mezclar las capas de aire, trayendo la más caliente a la superficie.
La energía radiada desde la superficie del suelo durante la noche es proporcionada en parte por el aire y en parte por las capas superiores del suelo.
Los ventiladores, al crear turbulencia, aumentan la conductividad del aire, (Budyko, 1974).
Las formas más frecuentes de humedecer la vegetación se da con el uso de sprays.
En el caso de suelos húmedos y zonas lacustres, el uso del spray es para producir una superficie monomolecular, impermeable al agua, eliminando efectos de evaporación; entonces al menos algo de la energía que se perdería como el calor latente, es disponible como calor sensible para calentar el suelo o el cuerpo de agua.
Esto es útil en el caso de cultivos donde la temperatura del agua es importante en el crecimiento de la planta, (Cooper, 1975).
La conservación del agua en presas es otra aplicación, pero existen problemas si el viento es mayor de 2 m s-1, ya que la capa aislante producida es removida.
Este sistema se ha utilizado también para disminuir la transpiración y evitar la pérdida de agua en la planta.
En algunos casos la intención es la de sellar el estoma, pero como la mayor parte de ellos se encuentra en la parte inferior de la hoja, hace muy difícil su aplicación.
Otros antitranspirantes operan induciendo químicamente a que los estomas se cierren o disminuyan su abertura incrementando así su resistencia estomatal.
Todos estos materiales deben de permitir el intercambio del dióxido de carbono, si se quiere que la planta se conserve sana.
Así como las cubiertas emplean las excelentes propiedades del aire, la irrigación además de reducir el potencial de stress de humedad del suelo, puede producir un clima del suelo más estable.
La adición de agua al suelo resulta en un incremento de la difusitividad térmica, y compensa valores extremos de calentamiento en el día y enfriamiento en la noche.
Con el incremento de disponibilidad de agua se incrementa la evaporación y se da un adicional efecto de enfriamiento.
El completo inundado de algunos cultivos es para utilizar las propiedades climáticas conservativas asociadas al agua.
Los principios de protección de heladas pueden observarse mejor en el balance energético nocturno.
El propósito esencial es de mantener la temperatura del sistema suelo-aire arriba de un valor crítico y evitar el daño a plantas sensibles.
Esto puede obtenerse en tres formas: a) Retardando la pérdida de energía; b) Redistribuyendo la energía existente dentro del sistema; c) Agregando nuevas formas de energía al sistema por medios artificiales.
Las medidas de protección son más efectivas en este último caso, ya que depende de propiedades locales y procesos que son más fácilmente controlables.
Es aconsejable evitar sitios donde el aire frío puede estancarse y acumularse.
Esto incluye áreas bajas como valles y depresiones de terreno y detrás de obstrucciones al flujo que baja en las laderas, como paredes, cercados, carreteras o terraplenes de ferrocarril.
Si tales barreras existen, el problema puede disminuirse con aberturas en las paredes o canales de desviación.
En valles grandes debe hacerse uso del "cinturón térmico" en lugar del fondo del valle.
Desafortunadamente la calidad del suelo es a menudo mejor en la .parte baja y el valor de esto tiene que ser pesado con el potencial de riesgo de helada.
La pérdida superficial de calor es la fuerza que determina el enfriamiento nocturno, tendiendo a predominar las heladas en noches sin nubes.
Un método de protección es el de tratar de evitar la pérdida de calor colocando una pantalla radiativa arriba de la superficie.
Con estas nubes de barrera, se absorbe gran parte de la energía de la superficie y se radiará una porción de regreso, por lo que las pérdidas de calor son correspondientemente reducidas.
Nubes artificiales de niebla y bruma (de sprays con agua) y humo (quemando llantas de carro) han sido usadas para obtener el control de la radiación.
Idealmente los materiales de las cubiertas deben ser aplicados en la tarde de una noche riesgosa de helada, esto permite el máximo almacenamiento de calor durante el día.
Las cubiertas señaladas anteriormente para retención de humedad y calor son útiles también para protección de heladas.
El segundo procedimiento es el de aumentar la conductividad térmica de la capa superior del suelo, así como maximizar la transmisión ascendente de calor en el suelo.
Esto puede hacerse agregando humedad a través de la irrigación o aplanando el suelo para extraer el contenido de aire, esta técnica es aceptable en el otoño, pero no puede repetirse frecuentemente.
Otro procedimiento es inundar completamente el suelo o el cultivo.
Ciertas plantas son capaces de resistir este tratamiento.
El balance energético será similar a un campo acolchonado que da un ambiente térmico más estable.
Las temperaturas a las que las plantas son dañadas, se da usualmente abajo de los 0°C, así que hay un pequeño rango de temperaturas dentro de las cuales el daño es sólo temporal.
El uso de sprays para protección de heladas, hace uso de este margen de seguridad y utiliza el hecho de que cuando el agua se congela libera calor.
Cuando plantas o partes de ésta se han enfriado ligeramente abajo de 0°C, se usa el spray fino de agua, conforme el agua se congela libera calor, ayudando a retardar el enfriamiento.
La adición de agua aumenta la capacidad calorífica de las plantas, retardando la razón de enfriamiento.
Dado que el uso del spray y el enfriamiento son continuos la temperatura estará próxima a 0°C sin ser dañado el cultivo.
El método requiere un control cuidadoso, si muy poca agua es agregada, la temperatura puede bajar por evaporación o sublimación, donde si se agrega mucho, la cantidad de calor liberado puede no ser suficiente para calentar la planta y equilibrar el enfriamiento radiativo y convectivo del exterior.
125
Si el spray no es usado el tiempo suficiente, el calor se perderá de la planta y el daño puede aparecer.
Durante las noches de heladas la atmósfera baja se caracteriza por una inversión (hay un considerable almacenamiento de calor en las capas superiores).
La base de la inversión está sobre el suelo desnudo y sobre el follaje (cuando hay vegetación), el mezclado vertical en combinación con esta distribución da lugar a un aumento de la entrada de calor a la superficie, este flujo puede incrementarse y la temperatura promedio de las capas bajas puede ser aumentada.
La cantidad de aumento de temperatura depende del grosor e intensidad de la inversión térmica.
ASPECTOS CLIMÁTICOS EN LA PRODUCTIVIDAD OCEÁNICA
Todo lo que existe sobre la superficie terrestre es constantemente influenciado por factores meteorológicos.
El motor generador de la energía que influye en nuestra vida diaria es el Sol, que desarrolla, anima y activa la, vida terrestre y marina, también es el .inhibidor y -responsable indirecto de su destrucción y muerte.
En el proceso de la vida, la radiación solar interviene en diversos niveles de producción: el radiativo, el termodinámico y el dinámico.
El efecto radiativo actúa directamente en el crecimiento de las plantas terrestres y fitoplancton marino a través de los procesos de producción fotosintética, cuya consecuencia es la generación de oxígeno que mantiene, tanto la vida terrestre como la marina, e indirectamente, en el nivel termodinámico, a través de la temperatura (relacionada con el almacenamiento de energía), que juega un papel importante en la fotosíntesis y en la cinética enzimática asociada a ella.
Las diferencias de temperatura, crean diferencias de densidad y presión, las cuales asociadas con la rotación de la Tierra -nivel dinámico- forman una circulación atmosférica y marina que intensifica o debilita a las corrientes marinas por diferencias de densidad y a los vientos por diferencias de presión.
Además, a través de los procesos de evaporación (se toma energía) y condensación (se libera energía), se transporta la mayor parte de la energía térmica de los océanos a la atmósfera donde los sistemas de vientos a gran escala realizan su transferencia de regiones con exceso de energía a otras con déficit (usualmente de regiones ecuatoriales hacia los polos), reforzando la circulación atmosférica (modificada por la forma de la Tierra, distribución de océanos y continentes, etc.) que influye en la dinámica de las masas de agua y sus movimientos horizontales y verticales los cuales transportan los nutrientes controlando éstos en gran medida la bioproductividad de los océanos.
La acción conjunta de estos tres niveles de influencia solar (radiativo, termodinámico y dinámico) favorece el desarrollo de la vida oceánica y cualquier situación anómala que presente cualquiera de ellos se hará notar en su productividad. (Ritter y Guzmán, 1978,1979).
La capacidad productiva resulta de dos procesos aparentemente antagónicos: la fijación fotosintética del dióxido de carbono y su liberación por respiración.
A través de la respiración se efectúa la oxidación de los componentes producidos en la fotosíntesis liberándose energía para la ejecución de los variados trabajos biológicos y químicos de la planta.
El Sol se encarga de abastecer la energía para la reacción fotosintética y los fermentos de orientarla en la dirección adecuada, es decir, las plantas tienen sus catalizadores, que son enzimáticos y cada enzima obliga a la reacción a que se desarrolle en un solo y determinado sentido.
En otros términos, las enzimas son específicas.
En el agua, cuando la radiación solar penetra se absorbe selectivamente y con mayor rapidez que en el aire.
Ocurriendo el mínimo de absorción en aproximadamente 0.47 m (azul), el ultravioleta (UV) es absorbido rápidamente, pero la principal absorción es al final del rojo del espectro, y en el infrarrojo (IR) aproximadamente el 90% de la radiación es absorbida en el primer metro.
De las sustancias disueltas en el agua, las más importantes absorben fuertemente en el violeta y el azul, moderadamente en las longitudes de onda media y débilmente a mayores longitudes de onda.
La mayor transmisitividad, se da cuando existen pequeñas cantidades de materia disuelta en el agua y es en el verde, y para grandes cantidades en el naranja y rojo.
La difusión y absorción se hacen más importantes a medida que aumenta el número de partículas en el agua.
La luz absorbida por una molécula sitúa a uno de sus electrones en un estado excitado de energía, quedando el electrón en disposición de aparearse con otro electrón de una molécula o átomo vecino en un enlace covalente.
Mediante este proceso fotoquímico se forman nuevas moléculas.
La luz que es absorbida por la molécula de clorofila y transferida a los electrones, actúa de tal manera que se crean fuertes oxidantes y reductores, es decir, moléculas que arrancan con facilidad electrones de otras (las oxidan) o los ceden (las reducen).
En términos de eficiencia de conversión de energía, las plantas son 100 veces más eficientes que cualquier máquina hecha por el hombre.
No toda la radiación solar sirve a los propósitos de ordenar la biosfera. En el ultravioleta UV la energía resulta excesiva (el electrón del átomo receptor se excita tanto que se escapa del átomo y éste se ioniza), en el infrarrojo IR la energía es insuficiente (el átomo que recibe la radiación vibrará más intensamente o rotará o se trasladará más rápidamente por su medio, pero la influencia que sufre no es lo suficientemente intensa para mover los electrones de sus órbitas), por lo tanto estas gamas de radiación solar no sirven para formar moléculas biológicas.
En cambio, las radiaciones de la zona visible tienen la energía adecuada (durante el tiempo que el átomo receptor está excitado, la probabilidad de asociarse a otro y formar una molécula más compleja, es mucho mayor que cuando no lo está), por lo tanto, la radiación visible constituye la fuente fundamental de energía de la biosfera. (Cerejido, 1978).
Al alumbrar las plantas con luz de diferente longitud de onda, éstas en el proceso de fotosíntesis forman compuestos de diferente naturaleza química.
Si las plantas se iluminan con rayos rojo-amarillos, los compuestos principales que se forman como resultado de la fotosíntesis serán hidratos de carbono.
Si los rayos son azules, se forman proteínas ( Vlasou, y. Trifonov, 1975).
Además, los rayos UV y azul-violeta retardan el crecimiento de las plantas, resultando que las plantas crecen más rápidamente en la noche.
La energía radiante almacenada en forma potencial por las plantas, sufre una lenta conversión a energía cinética que permite a las comunidades ecológicas sobrevivir.
Los componentes de las plantas serán convertidos en otros componentes en los herbívoros, y éstos en otros por los carnívoros y, al final, los descomponedores que completan el ciclo biogeoquímico regresan los nutrientes al ecosistema para ser usados nuevamente.
La energía solar que fluye a través del sistema no es destruida (primera ley de la termodinámica) y en cada nivel biológico de producción, una parte de ella es usada (10-20%) para soportar la vida y, una mayor cantidad es degradada de una forma de energía capaz de realizar reacciones y trabajo en formas menos útiles y regresa al ambiente como calor o entropía (80-90%), de acuerdo con la segunda ley de la termodinámica.
Entonces, podemos decir que dependemos en nuestra vida diaria de las cantidades de material orgánico, fundamentalmente proteínas, que pueden ser producidas por una cantidad finita de energía solar incidente sobre la superficie terrestre.
En varias áreas, la magnitud de cambio en la temperatura superficial, durante un periodo de pocos días puede igualar el rango total anual; esto sucede especialmente en latitudes bajas y medias, y en casos de zonas de surgencias; observándose los mayores cambios cuando varios factores actúan en el mismo sentido.
También la temperatura y la profundidad de la capa de mezclado pueden cambiar en pocos días más de 1°C y 30 metros respectivamente, especialmente si una tormenta pasa sobre el área.
El mecanismo más importante de cómo la temperatura influye en la supervivencia de las larvas es a través de la disponibilidad de alimento en el tiempo apropiado: temperaturas muy altas o muy bajas pueden poner el desarrollo de las larvas fuera de fase respecto a la máxima producción de alimento, ya que la producción fitoplanctónica está muy relacionada con los cambios estaciónales de temperatura y cantidad de luz.
Cuando la luz es limitante puede considerarse que el mecanismo de fotosíntesis es independiente de la temperatura; con luz fuerte, cuando los mecanismos no fotosintéticos son limitantes, parecen ocurrir fuertes efectos de la temperatura, (Rabinowitch, 1945).
La abundancia de zooplancton (importante como alimento para la larva), está relacionada a la abundancia de fitoplancton y al período de desove de animales planctónicos adultos, el cual también es controlado por la temperatura.
En latitudes medias y altas, las altas temperaturas coinciden con clases anuales pesqueras abundantes, mientras que las bajas temperaturas coinciden generalmente con clases anuales pobres.
Toda la evidencia sugiere además, que la razón de regeneración de nutrientes en las áreas costeras es dependiente de la temperatura: a mayor temperatura mayor será su razón de regeneración.
Esto está de acuerdo con el hecho de que la regeneración de nutrientes es debida principalmente a procesos microbiológicos y la intensidad del metabolismo bacteriano es dependiente de la temperatura.
- El sistema dinámico de remolino. Se produce por la colisión de dos sistemas diferentes de corrientes (frías y calientes) o pertenecientes a aguas oceánicas y aguas costeras.
Los centros fríos del remolino ciclónico, corresponden en el otoño a localidades de concentración de ballenas, mientras que en la primavera y principios del verano hay correspondencia con concentraciones de atún (albacora).
Los núcleos calientes de los giros anticiclónicos corresponden a mediados del verano a concentraciones de barrilete, albacora y ballenas.
El grado de enriquecimiento de estas regiones parece ser proporcional a la magnitud de vorticidad de los remolinos oceánicos o aproximadamente al gradiente de temperatura de las masas de agua en ambos lados del frente en desplazamiento.
- Sistema de remolinos topográficos u orográficos. Son remolinos que se forman alrededor de islas, .correspondiendo con lugares de concentración de calamares, macarela y pargos. En estas áreas topográficas están localizadas importantes pesquerías comerciales, con sistemas de remolinos estacionarios.
Se puede establecer, en general, que en alta mar el abastecimiento de nutrientes se realiza a través de las corrientes oceánicas, mientras que en las áreas costeras donde el agua de la zona eufótica está más o menos regularmente en contacto con el fondo, la descomposición microbiológica de materia orgánica es el factor más importante, además de que se regeneran sales conteniendo fósforo y nitrógeno.
MODELOS MICROMETEOROLOGICOS.
Todos los fenómenos que se producen en la superficie de la Tierra son manifestaciones energéticas, donde los procesos importantes para el hombre son unidireccionales porque van acompañados por una degradación energética que los hace irreversibles.
La energía solar, aunque existe en cantidades inmensas, fluye en forma diluida y su cantidad por volumen en cualquier momento es sólo de 6 X 1016 kcal/cm3 mientras que el carbón y el petróleo son tipos concentrados de energía potencial por volumen.
El proceso fotosintético concentra la energía solar diluida, obteniendo hasta 131kca1/m2 /día, en sistemas de tierras productivas, como los bosques y los campos cultivados durante sus períodos de máximo crecimiento.
La agricultura representa el 8.9% de la superficie fotosintéticamente activa del mundo, aproximándose al rendimiento máximo de la naturaleza, sin embargo su aportación anual a la producción de la tierra es mucho menor que la de los sistemas naturales, ya que sólo tienen hojas durante una parte del año.
Los modelos son representaciones de la naturaleza, con diferentes grados de abstracción, desde los de una sola ecuación, relacionando la transpiración de una vegetación a valores de radiación incidente, o modelos más complejos a resolver con programas de computadora.
El propósito es ensamblar el conocimiento físico micrometeorológico a la aplicación a problemas biológicos, simulando sus partes identificables, como el cambio atmosférico o vegetativo, conectadas en forma lógica y obteniendo respuestas por la ruta de la base física, rebasando la base empírica y estadística.
MODELO DE PROCESOS DE TRANSFERENCIA (CALOR, HUMEDAD) EN LA CAPA VEGETATIVA.- Se combina el concepto de balance de energía, resistencia aerodinámica, resistencia estomática y saturación del aire dentro de los estomas, para calcular la temperatura o evaporación de una hoja.
Podemos escribir el balance de energía unitaria en la superficie de la hoja como:
Q* = QH + LEv (1)
Podemos escribir el calor sensible, QH, en función de la diferencia entre la temperatura de la hoja a la temperatura del aire (θ= Th - Ta ) , y la resistencia al intercambio de calor sensible (r).
QH =
ρ Cρ(Th - Ta)/ r = ρ Cρ θ /r (2)Así también relacionando del flujo de vapor de agua a la diferencia de vapor de presión dentro y fuera de la hoja, considerando el de dentro saturado, (e* - e) = DVP, la razón de cambio de la presión de vapor de saturación con la temperatura (de*/dT), la constante psicosométrica (
ﻻl) y la suma de r y la resistencia estomática rs , podemos calcular el intercambio de calor latente como:LEv = ρ Cρ [ (DVP) + de*/dT(θ)]/ ﻻl (r + rs) (3)
Sustituyendo (3) y (2) en (1) y resolviendo, tenemos:
θ = (Q* - ρ Cρ (DVP)/
ﻻ1 (r + rs) ]/ (ρ Cρ/r + ρ Cρ/ de*/dT/ﻻ1 (r + rs) ) (4)Fórmula utilizable para estimar la razón metabólica de la hoja.
El cambio en el flujo de calor sensible con la altura depende del área foliar por unidad de volumen (d
/dz ) del exceso de temperatura de las hojas en relación al aire que lo rodea y efectos de ventilación.
La divergencia del flujo de calor sensible en el follaje, será dada como:
= ρ Cρ θ/r d
De igual forma la divergencia de calor latente es dada como:
L
= { ρ Cρ [ (DVP) + de*/dT (θ)] / ﻻl (r + rs) } d ζ/dz (6)
Estas ecuaciones (5) y (6) son la base de los modelos de intercambio de calor y humedad en un follaje.
El balance de energía del follaje a una altura z es dada por:
Q*(z) = QH (z) + LEv (z) QG
La densidad de flujo de calor sensible con la altura del follaje es dada como
QH(z) = -
ρ Cρ KH (z) (T/
z + r a)
Donde KH es la difusitividad del calor.
La densidad de flujo de calor latente con la altura del follaje es dada como
Donde Kv es la difusitividad para el vapor de agua y
q/
z es el gradiente vertical de la humedad específica sí;
Haciendo
tendremos que
Si Q*1 y Q*2 son la radiación medida en el tope y el suelo del follaje, tendremos
Donde;
Simplificando al hacer uso de las relaciones
y
Donde Tw es la temperatura del bulbo húmedo y e* (w) es el vapor de presión a la temperatura del bulbo húmedo.
Entonces
Despreciando la energía utilizada en procesos de fotosíntesis y respiración de la vegetación, podemos determinar los perfiles verticales de temperatura media y promedio de humedad absoluta del aire
, temperatura media y promedio de humedad absoluta en la superficie del follaje
.
Una vez que estos perfiles son conocidos es fácil determinar la densidad de los flujos verticales y la intensidad de las fuentes y sumideros de calor sensible y latente, así como la distribución vertical de la "Razón de Bowen del follaje",
Considerando la temperatura equivalente como:
Si los coeficientes de intercambio de calor y vapor de agua son iguales, tenemos la ecuación de balance para las diferentes alturas del follaje, como
o
También;
Donde
: es la temperatura media equivalente en la superficie del follaje, entonces,
151
Si
es fijo;
Los perfiles requeridos de fuentes y sumideros pueden ser inferidos conocidos
para las diferentes alturas.
La fuente de calor sensible por unidad de altura es dada como
BIBLIOGRAFÍA
ARNHEIM, R. (1971). Entropy and Art. An Essay on Disorder and Order. Berkeley: University of California Press.ASHBY, W. R. (1972). Introducción a la Cibernética,, Buenos Aires; Ediciones Nueva Visión.AYALA, F. C. (1987) La Naturaleza Inacabada. Biblioteca Científica Salvat, Barcelona, España. BARTENIEFF, Irmgard (1980). Body Movement: Copying with the Environment. New York: Gordon and Breach. BERTALANFFY Von, L. V. (1968) General System Theory, Brazilier, Nueva York. University of Maryland, USA.
BILLINGS, W. D. (1978). Plants and the Ecosystem. Wadsworth Publishing Company, U.S.A., 177 p. BILLINGS et al (1966). Photosynthesis and Respiration Rates of Rocky Mountain Alpine Plants under Field Conditions. Am. Midl. Nat. 75 (1). 34-44. BLUM, H. F. (1951). Times Arrow and Evolution. Jew Jersey: Princeton University Press. 9. BOLTZMANN Ludwing (1974). Theoretical Physics and Philosophical Problems. D. Reidel publishing company, Boston
- . BOTKIN, D. F'. (1969). Prediction of Net Photosyntesis of Trees from Light Intensity and Temperature, Ecology 50(5). 854-858.
BRILL, E. F. (1981). El Fin del Mundo por Hambre. Ed. Aura, 272 p.153BUDYKO, M. I. (1963). Atlas of Heat Balance of the Earth. Interdepartmenta1 Geophysica1 Comite under the Presidium of the Academy of Sciences of the U.S.S.R. Main Geophysycal Observatory. BUDYKO, M.I. (1974). Climate and life. Academic Press U.S.A., 508 p CARSTEN, B. (1987). La Vida, un estadío Intermedio. Biblioteca Científica Salvat. Barcelona España. CESARMAN, E. (1984). Hombre y Entropía. Ed. Gernika, 401 p.CESARMAM, F. (1972). Ecocidio. Estudio Psicoanalístico de la Destrucción del Medio Ambiente. México. Cuaderno de Joaquín Martín. CEREIJIDO, M. (1978). Orden, Equilibrio y Desequilibrio (Una Introducción a la Biología) Ed. Nueva Imagen. 224 pp, COOK, EARL. (1971). The Flow of Energy in an Industrial Society. Scientific American, September. COOPER, J. P. (1975). Photosyntnesis and Productivity in Different Environments. Cambridge Universitv Press. London. CHARNEY, J. G. (1963). "A Note on Large-Scale Motions in the Tropics". J. of Atmospheric Sciences. 20, 607-609. DUFIE, J. A. (1974) Solar Energy Thermal Processes. John Wiley and sons. London p. 386. EGLE and SCHENK. (1951). The Application of the Infrared gas Analyser in Photosynthesis research. En. Patterns of Primary Production in the Biosphere. HFH, Lieth, ed. Benchmark Papers in Ecology 8. U.S.A. 106-120. EMERY, F. E. (1969) Systems Thinking: Selected Readings, Penguin, Nueva York. 24. EINSTEIN A. (1952) Autobiographical Notes, Albert Einstein: Philosopher/Scientist,
.E STRADA-CAJIGAL V. (1985). Análisis Crítico de Procedimientos para estimar la radiación solar Proy. 3140. Inst. de Ing., UMAM 7-26 pp..FASHEUM, I. and M. D. DENNT. (1932). Interception of Radiation and Growth Efficiency in Field Beans (Vicia faba L.) Agric. Meteor., 26, 221-229. .FIFKIN, Jeremy. (1930). Entropy. New York: Viking. .FLOHN, H. (1967). "Dry Equational Zones and Asymmetry of the Global Atmosphenc Circulation". Bonner Meteor. Abahaoud, 7:3-8 p. .FORDHAM, D. P., Hood, L.M. and Malafant, K. W. (1997) Complexity in landscapes and resource planning: packaging science for decision makers. 1997 Proceedings of the Ecological Society of Australia, Chapter 22.
.FORRESTER, J. W. (1970) Industrial Dynamics. Cambridge, Mass., US: The MIT Press. .GATES, D. M. (1960). Energy Exchange in the Biosphere. Harper and Row. New York. 1
.GATES, D.M. (1965) . Energy, Plants and Ecology, Ecology 4ó (1.2) 1 -13. .GATES, D.M. (19 71)» The Flow of Energy in the Biosphere en; Energy and Power. Scientific American. Inc U.S.A. 43--52. .GEIGER, R. (1969) The Climate near the Ground. Harvard Univ. Press. Cambridge, Mass. .GRANT E. W., S. MARIN L. Y K. PEDERSEN E. (2001). Ecology and natural management:system analysis and simulation. John Wiley & sons, Inc. 36. HASSELL, M. P., J. H. LAWTON AND R. M. MAY (1976) patterns of dynamical behavior in single species populations. journal of animal ecology 45:471-486.
37. HOLLAND, J. H. (1995) Hidden Order: How adaptation breeds complexity, Addison-Wesley Publishing Co., Reading, MA.
38. HRABER, P. T. and Milne, B.T. (1997) Community assembly in a model ecosystem, Ecological Modeling, 103: 267-285.
- .JORGENSEN, S, Mejer, H. and Nielsen, S. (1998) Ecosystem as self-organizing critical systems, Ecological Modelling, 111: 261-268.
.ELMS, J.A. (1965). Diurnal and seasons patterns of net Assimilation in Douglas Fir. Pseudotsuga Menziessi (MIRB) Franco as influenced by environment, Ecology, 46, 698-708. .HUBELL S. (2001) The unified neutral theory of biodiversity and biogeography. Princeton .JACOB F. (2005) El juego de lo posible. Fondo de Cultura Económica, México, 96 pp. .JACOB F, (1986) La logica de lo viviente. Salvat Editores.44. JACOB F. E. (1980 ) The self-organizing Universe. New York Pergamon.
45. JACOBSON H. (1955). Information, Reproduction and the Origen of life. Amer. Sci. 43: 119-127.
Geoph. Biokl, Ser, B. 17. 413-428.
- .JENIK .J. (1969). Interpretation of Direct Solar irradiation in Ecology. Arch. Met.
.JONE:S, G. (1979). Vegetation Productivity, Longman Inc., U.S.A. 100 p. .KAHN, H, Y WIENER, A. J. (1967) The Year 2000, A. Framework for speculation the next; Thirty-Three Years. New York..KAUFFMANN, S., (1985). "Organization and Complexity". Oxford University Press. .KAUFFMANN, S. (1993) The origins of order: Self-organization and Selection in Evolution. Oxford University Press, New York. .KAUFFMANN, S. (1995) At Home in the Universe. Oxford University Press. New York .KIRA et al. (1967). Comparative Ecological Studies on Three main types of Forest Vegetation in Thailand. IV Dry Matter Production with Special Reference to the Fhao Chong Rain Forest en: Patterns of Primary Production in the Biosphere, H.S.G. Lieth, ed., Benchmark Papers in Ecology, 8 U.S.A., 125-153..KRAUS E.B. (1977). "Subtropical Droughts and Cross-equatorial Energy Transports" Mon. Wea. Rev., Vol. 105, 1009-1018, .KRUEGER, A..F. and T. I. GRAY Jr., (1969). "Longterm Variations in Equatorial Circulation and Rainfall" Mon. Wea. Rev., 97 (10): 700-711. MET-01-00014. .LANGTON, C. (1990) Computation at the edge of chaos: phase transitions and emergent computation, Physica D, 42: 12-37. .LAYSEN, D. (1975). The arrow of time. Scientific Amencan. 223(6): 56:;69. .LETAU, A. H. (1969) Evapotranspiration Climatonomy. Monthly Weather Rev. 97, 691-699. . LIETH And BOX (1972). Evapotranspiration and Primary Productivity C.W. Thornthwaite memorial model. In: Papers on Selected Topics in Climatology. J.R. Mather, ed. 2: 37-44, Elmer, New York. .LIETH, H., (1973) Primary Productivity in Ecosysteniss Comparative Analysis of Global Patterns. Unifying Concepts in Ecoloqy. pp. 67-68 .L ILIENFELD, Robert (1978) The rise of System Theory, Jhon Wiley, Nueva YorkLOTKA, A.J. (1925) Elements of Mathematical Biology. Williams and Wilkings, Baltimore. 61.
3. Animal. Ecol. 17: 75-80.
- .LOWRY, P.w. (1970) Weather and Lite. Academic Press. New York.
- .MARGALEF R. (1963a) On Certain Unifying Principies in Ecology.
.MARGALEF, R. (1968) Perspectives in ecological theory. Chicago: University of Chicago Press. .MARGALEF. R. (1980). La Biosfera entre la Termodinámica y el juego, Ediciones Omega. Barcelona, 236 p. . MARGALEF, R, (1981) Ecología. Planeta, España, 2a. Ed. , 252 p. 67. MARGULIS, Lynn (1998a) Symbiotic Planet, Basic Books, NY.
68. MAY, R. M. (1969). Estability and Complexity in Model Ecosys
tems. Princeton Univ. Press. N.J.69. MAY, R. M. (1972a) On relationships among various types of population models. Amer. Nature, 107, 46-57.
70. MAY, R. M. (1972b) Will a large complex system be stable?. Nature, 238, 413-414.
71. MAY R. M.,and G.F. Oster(1976) Bifurcations and dynamic complexity in simple ecological models, Amer. Natur. 110(1976), 573-599.
72. MAY, R. M. (1976) Simple mathematical models with very complicated dynamics. Nature 261:459-467.
Agric . Meteor. 2O, 221-229.
- .MICHALCZYK, K.W, (1979). The Effect of Climatic Variatlons an the Significance of Agricultural Planning data.
.MILANKOVITCH, M. (1920). Theorie Mathematique des phénomenes thermiques produits par la radiation solaire. Gauthler Villiers et cie. Editeur. Paris..MILLER, P.C. (1970) Sampling to Estimate mean leaf Tenmperatures and Transpiration Rates in Vegetation Canopies. Ecology, 52 (5). 885-889. 76. MINGERS, J.,(1995) Self-Producing Systems.
Plenum, Nueva York.Chance and Necessity. New York, Knopf. 77. MONOD, Jacques (1971).
78. MONTEITH. J.L. (1965a) Radiation and Crops. Exp. Agric. Rev., 1, 241-51.
79. MONTEITH, J.L. (1975, 1976) Vegetation and Atmosphere,Vols 1 and 2. Academic Press, London.
.MORENO P. M. A. RITTER O.,W. REYES J.M.. (1989). Microfísica de Cultivos y Sistemas Naturales. Univ. Aut. de Gro..MORGEN, A. (1957) Die Besonnung und ihre Verminderung durch Horizontsont begrenzug. Veroff, Meteor. Hidrol. Dienstes AAR 12, 1-16.MUNN, R.E. (1970). Biometeorological Methods. Academic Press. New York. 366 p. .NAMIAS, J. (1963) "Response of the Eguatorial Counter Current to the Subtropical Atmosphere", Science, 28 sept. 181: 1244-1245. P/0-01-00005. 84. NICOLIS N., and Prigogine I. (1977) Self-organization in non-equilibrium systems, Nueva York, Wiley.
. ODUM, P.E. (1971). Ecología, Ed. Interamericana, México, 3a,. Ed., 639 pp..ODUM And PINKERTON (1955). " Time`s Speed Regulator: The Optimum Efficiency for Maximun power output in physical and Biological Systems" Amer. Scientist. 43, 331-343.15
.OKE, T.R. (1978). Boundary Layer Climates. John wiley and Sons. New York. 372 p. 88. PARSEGIAN, V. L. (1972).
The Cybernetic world. Double day & Company Inc. Garden City, New York.89.
PATRON, S. R. Y JONES. J.G. (1989). Soyabean production in the tropics. A simulation case for México. A simulation case for México. Agricultural Systems 29. 219-231.90. PIRIE, N.W. (1969) Food Resources Conventional and Novel Penguin Books, Baltimore Maryland, 208 p.
91. PRIGOGINE, I., (1980) From Being to Becoming, Freeman, San Francisco.
92. PRIGOGINE, I., and Stengers I. (1984) Order out of chaos, New York, Bantam.
93. PRIGOGINE, I., (1989). The philosophy of instability. Future, 21, 4:396460.
94. PRIGOGINE, I., (1997) The End of Certainty. Taurus, Madrid.
95. QUASTLER, E. (1964). The emergence of Biological Organization (New Haven, Conn: Yale University Press).
96. RABINNOWITCH E. I. (1945) Photosynthesis and Related Processes". New York, Interaciencie Publishers.
97. REITER, E. (1977a). "The Interannual Variabi1ity os the Qcean-Atmosphere System". J. Atmos. Sci., 35(3), 349-370.
- .REITER, E. (1977b). "Long-Term Variabi1ity in the Tropical Pacific its Possible Causes and Effects" Mon. Wea. Rev., 106, 324-330.
.RITTER, O.W., Y GUZMAN, R. S. (1978). Efectos meteorológicos en la Producción Oceánica del Golfo de Panamá, Meteorológica, Vol. VIII/IX, Págs. 349-358.100. RITTER. Q. W., Y GUZMAN, R. S. (1979). Resultados Preliminares del Estudio de los efectos meteorológicos en la productividad oceánica del Golfo de
Panamá Geofísica Internacional.
101. RITTER, O. W., GUZMÁN, S. (1982.)
*Efectos meteorológicos y oceanográficos en la productividad del Océano Pacífico del Este*. Revista de Geofisica i.p.g.h.(oea.) vol.17, págs. 23-39.102. RITTER, O. W., GUZMÁN, S. (1984.)
*Modelo generalizado de producción pesquera con dependencia ambiental: una aplicación al Golfo de Tehuantepec*. Revista Geofísica i.p.g.h. (OEA). vol.20, págs. 21-29.103. RITTER et al (1986).
Balances de radiación y potenciales de Productividad primaria en algunos ecosistemas terrestres de la República Mexicana. Geofís. Int. 25, 2, 285- 314.104. RITTER et al (1987). Efectos Estocástico-Ambientales en el Crecimiento poblacional atunero en el Océano Pacifico del Este. Meteorológica Argentina).(Vol. XV, No. 1).
- . RITTER et al (1988),. Evaluación del Potencial y la Estabilidad de la Producción Agrícola con base en índices Climáticos para algunas localidades de la República Mexicana. Geofísica Internacional
106. RITTER,O W., SUÁREZ, R. RODRÍGUEZ (1992.) Crecimiento, sobrevivencia y optimización de la carpa (cyprinus carpio) en la presa de Atlangatepec, Tlaxcala. Anales del Instituto de Ciencias del Mar y LImnologia, UNAM vol. 19, no.1, págs.
(43-56).-107. RITTER, O. W., KLIMEK, R., PATIÑO, R. (1997.).
Sobre el pronóstico en los sistemas fisico-climaticos y biológico-productivos, posibilidades y limitaciones. revista geografica del i.p.g.h., (OEA), núm. 123, pp. 203-218.108. RITTER, O.W., Mosiño, P. A., Patiño R. M., (2000a) "Pronóstico y naturaleza". Revista Ciencia y Desarrollo. julio/agosto del 2000.
volúmen XXVI, no. 153.
109. ROSEN, R. (1970) Dynamical system theory in biology. vol. 1. New York: John Wiley and sons.
110.
ROSENBERG, N.J. (1974). Microclimates The Biological Environment, Jonn Wiley &: Sons. New York.111. ROSENTHAL, S.(1967). "Some Preiliminary Theoretical Considerations of Tropospheric Wave Motions in Equatorial Latitudes". Mon. Wea. Rev., 93. 607-612.
112. ROSENZWEIG, M.L. (1968). Net Primary Productivity of Terrestrial communities: Prediction from Climatological Data. Am. Nat. 102 (923), 67-74.
- . SAMOYLENKO, V.S. (1970). "The Effect of wind Solar Radiation upon the Qcean" (Nature of the Perú Current). Oceanology Vol. 10(1): 1-12 Transl. A.G.U. from Oceanology, Academy of Science. URSS-MET-02-00013.
- . SAWYER, J.S. (1964). "Notes on the Possible Physical Causes of Long Term weather Anomalies " . Symposium Long Range Forescasting. Boulder, Colorado. Technical Note No. 66, WMO 162, T.P. 79, 227-248p. MET-03-00013.
- . SCHRODINGER, E., (1945). what is life? Cambridge, England University Press. Chicago.
- . SELLER, W.D. (1965). Physical Climatology. Univ. Chicago Press, Chicago.
- . SHANNON, C.E. And W. Weaver (1949). The Mathematical Theory of Communication. Urbana Univ. of Illinois Press.
- . SLOBODKIN, L. B. (1960). Ecological Energy Relationships at the Population Level. The American Naturalist, XCIV: 213-236.
119. SOLE, R. V. AND BASCOMPTE, J. (1998) Emergent phenomena in spatially extended model ecosystems. In modeling Spatiotemporal Dynamics in Ecology, (eds.) R. V. Sole and J. Bascompte, 1-25, Springer-Verlag, Berlin.
- .SNEVA, F.A. (1982). Relation os Precipitation and Temperature with Yield of Herbaceous Plants in Eastern Oregon. Int. J. Biometeor., 26(4), 263-276.
- .STANHILL et al (1965). Radiation Balance of Natural and Agricultural Vegetation, Quart. J. Roy. Met. Soc.. 92, 128-140.
122. STEELE, J.H. (1965). Notes on Some Theoretical Problems in Production Ecology. ín "Primary Productivity in Aguatic Environments" (C.R. Goldman, Ed.) pp. 385-398. Mem. Ist. Ital., Hidrobiol., 18 suppl., Univ.
Calif. Press Berkeley
. STEVENS, P.S. (1989). Patrones y Pautas en la Naturaleza. Biblioteca Científica Salvat, Barcelona, España.124 .SUTTON, O.G. (1953).
Micrometeorology. Me Graw Hill. New York.125 .THORNWAITE. C.W. (1951). The Water Balance in Tropical Climates. Bull Am. Meteor. Soc. 32(5), 166-173.
- .THORNWAITE and MATHER. (1957). Instructions and Tables for Computing Potential Evapotranpiration and the water balance. Centeton. N. Jersey. U.S.A. 311 pp.
- . UDA, M. and I. MAKOTO. (1958). "Enrichment Pattern Resulting from Eddy System in Relation to Fishing Grounds". J. of the Tokio University of Fisheries, 44 (1.2).
158
. VITREVIVH. (1960). Agricultural Meteorology, National Science Foundation, Jerusalem, 320, p. . VLASOV, I. and TRIFONOV, D. (1975) "Química Recreativa". Divulgación Científica (4), Ediciones de Cultura Popular, 341, pp. 130 .
VOCUM et al. (1964). Photosynthesis under Field Conditions. VI Solar Radiation Balance and Photosynthesis Efficiency. Agronomy Journal, 56(3), 249-253.131. VOLLENWEIDER, R.A. (1970). "Models for Calculating Integral Photosynthetic Productivity" pp. 455-472. Proc. IBP/PP Technical Meeting, Trebon 14-21 September 1969.
132. WIEGER, R. G., OWEN, D. F.(1971) Trophic structure, available resources and population density in terrestrial versus aquatic ecosystems. J. Theor. Biol. 30, 69-89.
133. WIELGOLASKI, F.W. (1975). Productivity of Tundra Ecosystems. En:Patterns of Primary Production in the Biosphere. H.F.H. Lieth, Ed., Benchmark Papers in Ecology, B, U.S.A.
134. WIENER, M., (1948) Cybernetics. New York, Wiley.
(Volver a página inicial)